Причины и механизмы развития аллергического ринита
Этиология аллергического ринита комплексная, сочетающая в себе генетические факторы и факторы окружающей среды. Аллергический ринит передается по аутосомно-доминантному типу с неполной пенетрантностью. Гены главного комплекса гистосовместимости (ГКГС), расположенные на шестой хромосоме, вызывают повышенный уровень продукции IgE В-лимфоцитами после контакта с аллергеном. IgE вызывает развитие аллергической реакции. Провоцирующий фактор для повышения экспрессии данных генов пока не обнаружен.
Аллергическая реакция представляет собой сложную последовательность событий с участием многих типов клеток, медиаторов и продуктов клеточной жизнедеятельности. Наши знания об аллергическом рините получены из двух основных источников: экспериментов in vitro; и исследований, в которых сначала человека подвергают воздействию аллергена, а затем анализируют клеточный состав смывов из полости носа и биоптата слизистой.
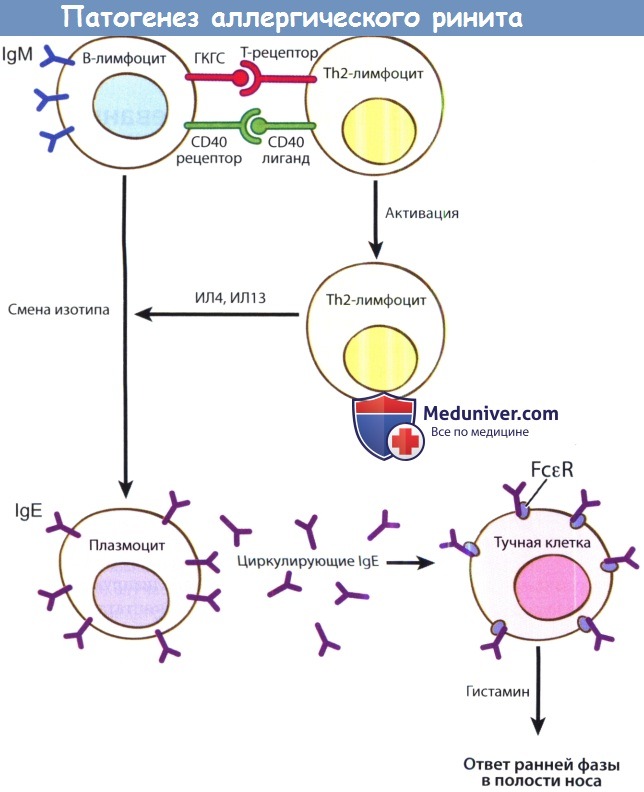
Все начинается с процессинга антигена, включающего дендровидные клетки, В-лимфоциты и макрофаги. В лимфатических узлах антиген представляется Т-хелперам второго типа CD4+ (CD4+ Th2-лимфоцитам). Затем В-лимфоциты продуцируют антиген-специфические IgM, a после получения сигнала от CD4+ Th2-лимфоцитов начинают продукцию Ig Е.
Аллерген-специфический IgE затем присоединяется к высокоафинным рецепторам, Fc-эпсилон (FceR), на поверхности тучных клеток и базофилов. Присоединение аллерген-специфических IgE к Fce-рецепторам вызывает дегрануляцию базофилов и тучных клеток, что и вызывает повышение клеточной активности и развитие воспалительных реакций, свойственных людям с аллергическим ринитом.
Более подробно этот сложный механизм, наше представление о котором постоянно развивается, представлен ниже. Более подробную информацию можно найти в руководствах по иммунологии.
Патофизиология аллергического ринита.
FceR — Fc-эпсилон рецептор; Ig — иммуноглобулин; IL—интерлейкин;
МНС—главный комплекс гистосовместимости; TCR-рецептор Т-лимфоцита;
Th2-лимфоцит CD4+ Т-хелпер второго типа.
Процессинг и презентация антигена. Реакция начинается с захвата и процессинга антигена В-лимфоцитами респираторного эпителия. Затем антигены обрабатываются ГКГСII класса и транспортируются на клеточную мембрану, где происходит их презентации CD4+ Th2-лимфоцитам. Для начала продукции базофилами IgM необходимо два сигнала. Первым является распознавание комплекса аллерген-ГКГС CD4+ Тh2 - лимфоцитами при помощи рецептора Т-клетки.
Затем происходит взаимодействие CD40-лиганда Th2-лимфоцитов с CD40-рецепторов В-лимфоцитов, которое и заставляет В-лимфоциты продуцировать аллерген-специфический Ig М. Активация CD-4 Th2-лимфоцитов вызывает секрецию цитокина IL-4. Интерлейкин-4 имеет множество функций, но одной из главных является смена секреции IgM на IgE в В-лимфоцитах. Затем В-лимфоциты созревают в долгоживующие плазмоциты, отвечающие за продукцию аллерген-специфических IgЕ.
Стимуляция базофилов происходит после того, как преформированные циркулирующие аллерген-специфические IgE соединяются с высокоаффинными FcεR-рецепторами на базофилах и тучных клетках. При присоединении свободного аллергена к аллерген-специфическому комплексу IgE- FcεR происходит перекрестная реакция, в результате которой происходит дегрануляция медиаторов воспаления, главным образом гистамина, из тучных клеток или базофилов, а также активируется фосфолипаза А2 на клеточной мембране.
Базофильные клетки высвобождают в эпителий ряд химических веществ, среди которых экзогюкозидазы, фактор некроза опухолей а (ФНО-а), гранулоцит-ассоциированные триптазы (только у тучных клеток), химаза, пероксидаза и арильсульфатаза В. Эти вещества вызывают повышение проницаемости сосудистой стенки, вазодилятацию сосудов слизистой оболочки полости носа и непосредственную стимуляцию нервных окончаний, что приводит к развитию симптомов ринита: зуда, чихания, ринореи, затруднения носового дыхания. Гистамин является основным медиаторов этого этапа, который еще называют «ответом быстрой фазы».
Взаимодействие аллергена с FcεR-рецепторами также вызывает транскрипцию цитокинов и инициацию распада арахидоновой кислоты. Секреция цитокинов и хемокинов усиливает развитие аллергической реакции. Продукция вазоактивных агентов и цитокинов, в числе которых ИЛ-1, ИЛ-2, ИЛ-3, ИЛ-4, ИЛ-5, ИЛ-6, гранулоцитарно-моноцитарный колониестимулирующий фактор (ГМКФ) и ФНО-а вызывают хемотаксис, клеточную пролиферацию, продукцию IgE и инфильтрацию слизистой оболочки носа, усугубляя проявления аллергической реакции.
Клеточный состав представлен эозинофилами, Т- и В-лимфоцитами, тучными клетками. ИЛ-5 отвечает за пролиферацию и выживание эозинофилов, а также за хемотаксис в слизистой оболочке носа. В результате дегрануляции эозинофилов высвобождаются высокотоксичные белки и кислородные радикалы, повреждающие структуру эпителия полости носа.
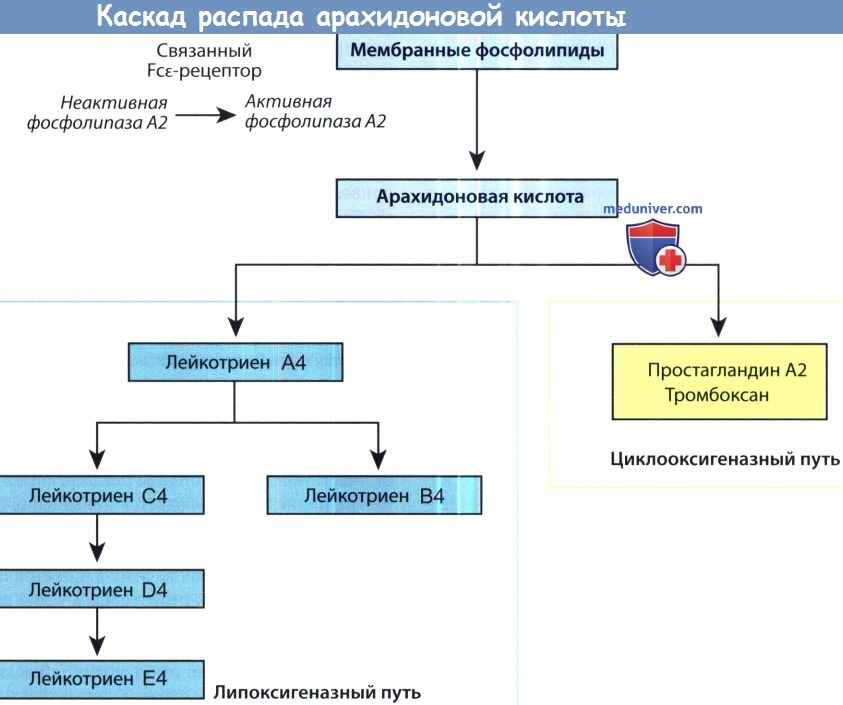
Каскад распада арахидоновой кислоты.
FcεR, Fc-эпсилон рецептор.
Метаболиты арахидоновой кислоты являются мощными медиаторами аллергической реакции. Активация фосфолипазы А2, следующая за активацией FcεR-рецепторов, приводит к высвобождению арахидоновой кислоты из клеточной мембраны. Метаболизм арахидоновой кислоты может протекать по двум путям: по циклооксигеназному с формированием простагландина А2, или по тромбоксановому или липооксигеназному пути с формированием лейкотриенов. Продукты распада арахидоновой кислоты вызывают затруднение носового дыхания, нейтрофильную инфильтрацию, вазодилятацию и клеточный хемотаксис.
Примерно у половины больных с аллергическим ринитом в течение 4-6 часов развивается ответ поздней фазы воспаления. По нашему мнению, за его инициацию отвечают базофилы, т. к. при анализе смывов полости носа у таких больных выявляется наличие тех же медиаторов воспаления, которые свойственны первой фазе, за исключением простагландина D2, в синтезе которого как раз не участвуют базофилы. В то же время, базофилы присутствуют в смывах из полости носа у больных с аллергическим ринитом.
Другими двумя важными факторами, участвующими в формировании симптомов аллергического ринита, являются прайминг-эффект и назальная гиперреактивность. Прайминг-эффект возникает при длительном контакте с аллергеном, ведущим к развитию хронических воспалительных изменений. Дозы аллергена, необходимые для развития симптомов аллергического ринита, постепенно уменьшаются. Хронические воспалительные изменения также ведут к повышенной реактивности, когда даже неспецифические ирританты, такие как сильные запахи, дым или примеси вызывают симптомы ринита.
Факторы окружающей среды также играют важную роль в развитии аллергического ринита. Особого интереса заслуживают активируемые протеазами рецепторы. Всего существует четыре вида таких рецепторов, из них 2 и 4 участвуют в формировании аллергических реакций. Эти рецепторы представляют собой связанные с G-белками трансмембранные рецепторы, которые экспрессируются многими типами клеток, в том числе респираторным эпителием полости носа. Стимуляция этих рецепторов необратима и вызывается эндогенными и экзогенными сериновыми протеазами.
К эндогенным протеазам относится триптаза, продукт дегрануляции тучных клеток и эозинофилов; экзогенные протеазы вырабатывают пылевые клещи, тараканы, альтернарии и аспергиллы. Стимуляция данных типов рецепторов вызывает повышение продукции IgE, лейкоцитарную инфильтрацию, рубцевание и развитие гиперчувствительности дыхательных путей. Окончательным эффектом стимуляции этих рецепторов является проникновение бытовых аллергенов в слизистую оболочку полости носа. Контакт с экзогенными протеазами в детском возрасте может быть фактором риска для развития аллергического ринита.
Роль цитокинов в иммунном ответе:
• Интерлейкин-2: активация Т- и В-лимфоцитов.
• Интерлейкин-4: аутопролиферация Т-лимфоцитов, активация В-лимфоцитов, рекрутинг клеток воспаления.
• Интерлейкин-5: рекрутинг и пролиферация эозинофилов.
• Интерлейкин-13: рекрутинг и пролиферация эозинофилов.