Как и в развитии многих структур глаза, Pax6 играет ключевую роль в развитии хрусталика. Первоначально Pax6 экспрессируется эктодермой головы, но затем под действием сигналов TGFβ нижележащих мигрирующих клеток нервного гребня его экспрессия ограничивается презумптивной плакодой хрусталика.
Нарушение экспрессии Pax6 у Xenopus вызывает формирование в глазном пузыре эктопического хрусталика. Сигналы Pax6, принадлежащего к семейству факторов транскрипции Meis, Bmp7 и Fgf определяют региональную локализацию плакоды еще до утолщения эктодермы. И у Bmp4-null, и у Bmp7-null мышей не происходит индукции развития хрусталика.
Разрыв сигнальной системы Fgf приводит к редукции экспрессии Pax6 плакоды и нарушает раннее развитие хрусталика и инвагинацию хрусталиковой ямки. Для ранней экспрессии гена кристаллина хрусталика необходимо связывание Sox2 (или Sox1) и Pax6 с энхансерами. Ген FoxE3 из семейства генов forkhead является геном-мишенью Рахб, он экспрессируется в презумптивной хрусталиковой плакоде.
Мутации FoxE3 у человека и мышей вызывают афакию и микрофтальмию. При мутациях Mab21l1 в хрусталиковой плакоде не происходит экспрессии FoxE3, даже несмотря на то, что Рахб сохранен; это указывает на то, что активация Mab21l1 происходит между активацией плакодного Pax6 и активацией FoxE3. У мышей с дефицитом Mab21l1 вследствие неполной инвагинации плакоды из-за недостаточной экспрессии FoxE3 при нормальной экспрессии Pax6, Sox2 и Six3 развиваются рудиментарные хрусталики. Семейство факторов транскрипции типа лейциновой застежки (basic leucine zipper transcription factors) MAF также участвует в развитии хрусталика и регулируется Pax6.
После отделения хрусталикового пузырька от поверхностной эктодермы важное значение приобретает экспрессия Prox1 для дифференцировки и элонгации клеток хрусталиковых волокон; при мутации Proxl не происходит поляризаии и элонгаии хрусталика, что приводит к развитию полого хрусталика. Экспрессия Pax6 в эпителии хрусталика прекращается чуть позже закрытия полости, тогда как Sox2 подавляется вскоре после образования полости. Sox1 экспрессируется на протяжении всего этого периода, он заменяет Sox2 и присутствует в клетках первичных волокон. Мутации Sox1 у мышей вызывают катаракту.
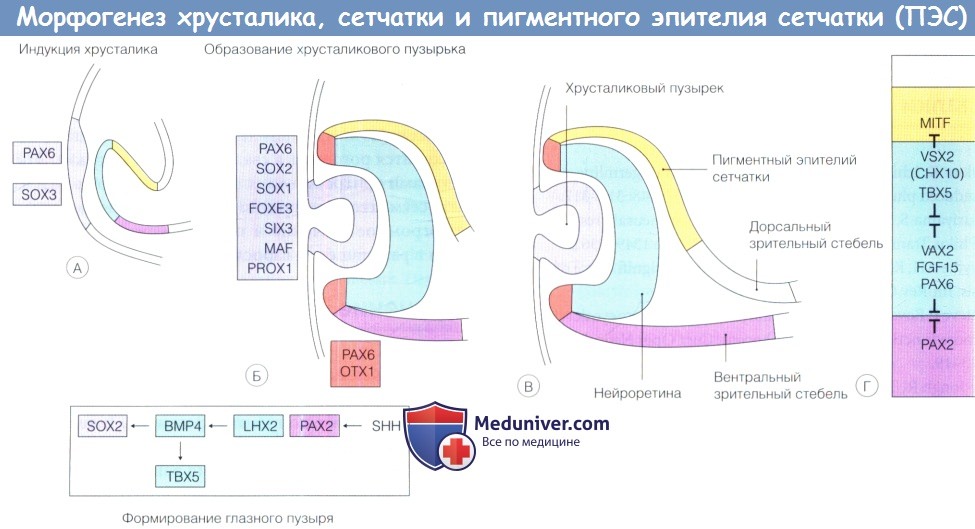
Формирование хрусталика, сетчатки и пигментного эпителия сетчатки (ПЭС):
(А) Для формирования хрусталика и образования хрусталиковой плакоды требуется сигнальная система между поверхностной эктодермой и глазным пузырем.
Формирование зрительного стебля происходит под влиянием индукции РАХ2 посредством SHH.
В презумптивной нейроретине LHX2 индуцирует экспрессию ВМР4, который, в свою очередь, индуцирует экспрессию SOX2 в поверхностной эктодерме; это запускает каскад экспрессии факторов трансдукции в хрусталиковой плакоде и индуцирует образование хрусталикового пузырька.
(Б, В). ВМР4 также индуцирует экспрессию ТВХ5.
(Г) Сложная сеть факторов транскрипции посредством индукции и репрессии определяет границы между зрительным стеблем и презумптивной нейроретиной, презумптивной нейроретиной и пигментного эпителия сетчатки (ПЭС).