Для дифференцировки мультипотентных прогениторов сетчатки необходимо взаимодействие активирующих и подавляющих факторов транскрипции bHLH (спираль-петля-спираль — basic helix-loop-helix, bHLH), так как для каждого типа клеток сетчатки характерен специфический код комбинации факторов bHLH.
Hes1 и Hes5 являются подавляющими bHLH молекулами, и у двойных нуль-мутантов глазные пузыри отсутствуют. Ганглиозные клетки сетчатки развиваются из прогениторов, коэкспрессирующих Pax6 и ген-активатор bHLH atoh7. Sox2 интенсивно экспрессируется в прогениторах нейронов.
Гетерозиготные мутации потери функции Sox2 вызывают двустороннюю анофтальмию и тяжелую микрофтальмию. Гипоморфные мутации Sox2 приводят к отсутствию ганглиозных клеток и вызывают нарушение стратификации нейроретины. Снижение уровня Sox2 сопровождается снижением экспрессии Notch1 и Hes-5, тогда как экспрессия atoh7 и Pax6 усиливается. Эти данные подтверждают ключевую роль Sox2 в развитии прогениторов нервных элементов сетчатки.
Shh действует как митоген клеток-предшественников сетчатки, под его влиянием увеличивается количество нервных клеток в экспериментах с культурами клеток. Рецептор Shh, Patched, экспрессируется в нейробластном слое сетчатки, характер его экспрессии следует за дифференцировкой ганглиозных клеток, сначала он экпрессируется в слое ганглиозных клеток, затем во внутреннем ядерном слое.
Активаторы bHLH — Neurodl и Neurod4 — экспрессируются в дифференцирующихся амакринных клетках. У мышей двойных мутантов амакринные клетки полностью отсутствуют, а число ганглиозных клеток и глиальных клеток Мюллера увеличено. Коэкспрессия Рахб/Neurod1 или Pax6/Neurod1/Neurod4 стимулирует дифференцировку амакринных клеток, а при коэкспрессии Pax6/Neurod4 горизонтальных клеток появляется больше, чем амакринных. Биполярные клетки одновременно экспрессируют Vsx2/Neurod4/Ascl1 (другой активатор bHLH).
Горизонтальным клеткам требуется коэкспрессия Pax6/Neurod4/Proxl/Foxn4. Для развития палочек и колбочек необходима коэкспрессия Neurod1, Ascl1, Crx и Otx2. Crx необходим для нормального функционирования палочек и колбочек, у человека он участвует в патогенезе дегенерации фоторецепторов и врожденного амавроза Лебера (Leber’s congenital amaurosis—LCA; MIM#602225).
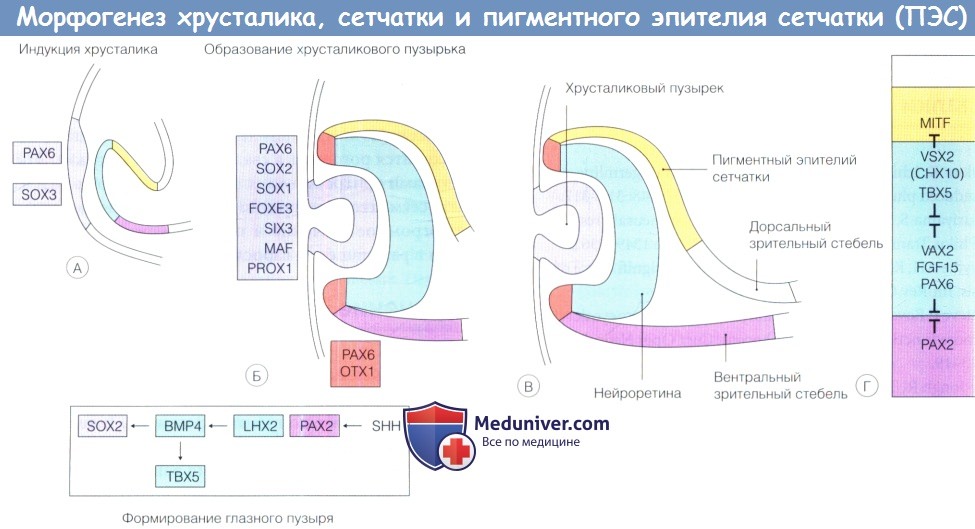
Формирование хрусталика, сетчатки и пигментного эпителия сетчатки (ПЭС):
(А) Для формирования хрусталика и образования хрусталиковой плакоды требуется сигнальная система между поверхностной эктодермой и глазным пузырем.
Формирование зрительного стебля происходит под влиянием индукции РАХ2 посредством SHH.
В презумптивной нейроретине LHX2 индуцирует экспрессию ВМР4, который, в свою очередь, индуцирует экспрессию SOX2 в поверхностной эктодерме; это запускает каскад экспрессии факторов трансдукции в хрусталиковой плакоде и индуцирует образование хрусталикового пузырька.
(Б, В). ВМР4 также индуцирует экспрессию ТВХ5.
(Г) Сложная сеть факторов транскрипции посредством индукции и репрессии определяет границы между зрительным стеблем и презумптивной нейроретиной, презумптивной нейроретиной и пигментного эпителия сетчатки (ПЭС).