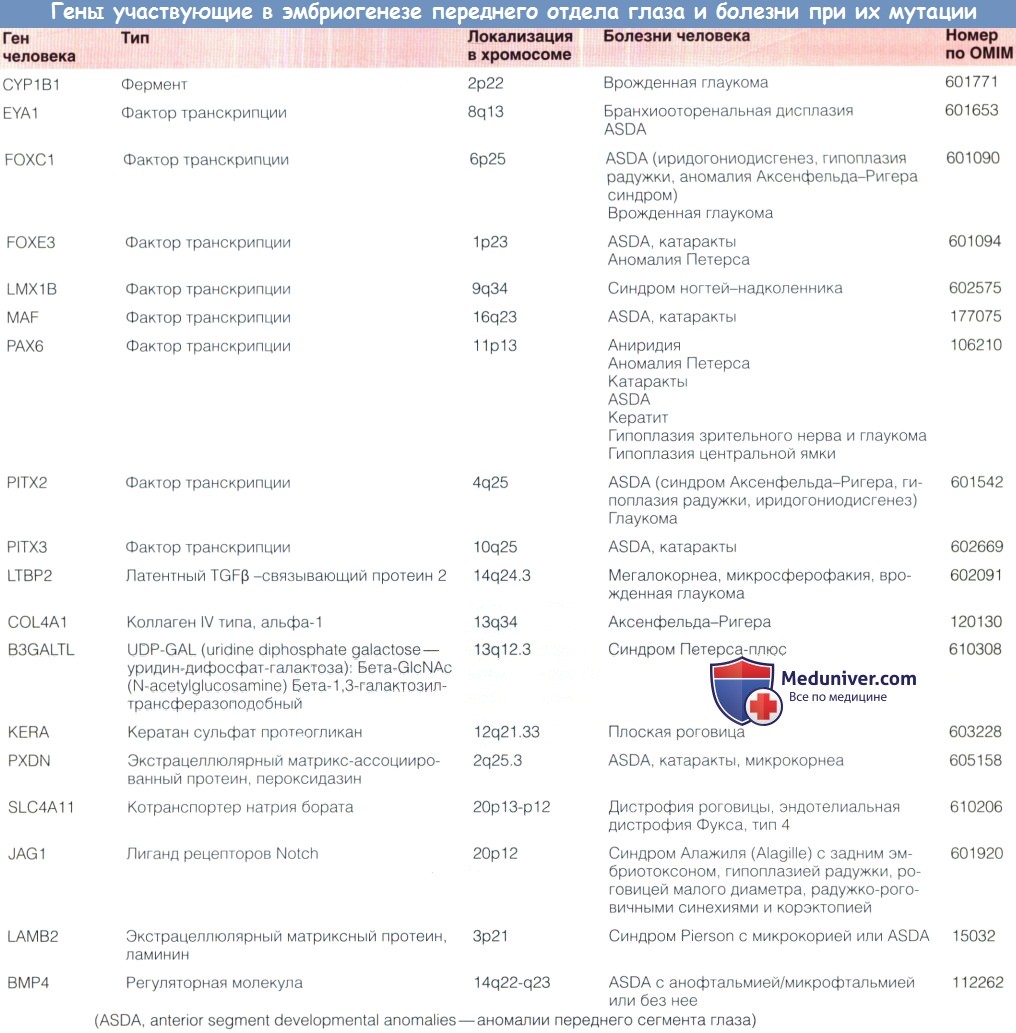
Гены контролирующие эмбриогенез переднего сегмента глаза
а) Мутации генов, вызывающие аномалии развития переднего сегмента. Было выявлено несколько заболеваний, вызываемых изменениями генов, на моделях животных была показана важная роль этих генов в процессе нормального развития переднего сегмента на молекулярном и клеточном уровне.
б) Факторы транскрипции и развитие переднего сегмента. Большинство известных на данный момент генов, играющих ключевую роль в развитии аномалий переднего сегмента глаза (anterior segment developmental anomalies — ASDAs), кодируют факторы транскрипции, которые регулируют транскрипцию других, целевых нижележащих в каскаде, генов.
Усиливая или подавляя экспрессию своих целевых генов, они координируют сценарии роста и дифференцировки. Каждый фактор транскрипции имеет свой тип ДНК-связывающего домена, который взаимодействует с регуляторной ДНК-последовательностью целевого гена. Эти ДНК-связывающие последовательности высоко консервативны в пределах царства животных, и отчасти поэтому животная модель оказалась полезной для понимания функции генов человека.
РАХ6 и PITX2 кодируют протеины, содержащие гомеодомены типов paired и bicoid, соответственно, тогда как FOXC1 кодирует протеин, содержащий домен forkhead. Эти гены очень близки генам, названным paired, bicoid и forkhead, функция которых была впервые определена в процессах развития и структурирования тела Drosophila melanogaster, которая является бесценной моделью для исследования многих генов, играющих роль в развитии человека и возникновении заболеваний.
в) Экспрессия генов развивающегося переднего сегмента: зоны действия генов. Зоны экспрессии генов во время развития переднего отрезка указывают на область их действия. Данные о зонах экспрессии генов получены в основном при изучении мышей как модели развития млекопитающего, также ограниченные сведения об экспрессии генов получены в результате изучения человека и других животных моделей и экспериментальных систем. Эта информация помогает понять причины развития аномалий, наблюдаемых у пациентов с генными мутациями.
Профиль экспрессии генов, мутации которых вызывают аномалии развития переднего сегмента можно разделить на три типа:
1. Экспрессируемые мигрирующими периокулярными клетками нервного гребня (Foxc1, Pitx2, Lmx1b)
2. Экспрессируемые только развивающимся хрусталиком (Foxe3, Maf)
3. С более обширной экспрессией во всех тканях глаза, в том числе в хрусталике, единственным примером такого гена является Рах6.
Рах6 экспрессируется на самых ранних стадиях развития глаза, этот ген получил название мастер-ген глаза (eye master control gene). Экспрессия Рах6 продолжается в клетках — производных нейроэктодермы глазного бокала и передней поверхностной эктодермы в течение всего развития глаза. Рахб экспрессируется в развивающемся хрусталике, внутреннем и пигментированном слоях радужки и цилиарного тела, эпителии роговицы и развивающейся сетчатке. В значительной степени он экспрессируется позже в развивающейся из мезенхимы трабекулярной сети.
В отличие от широко экспрессируемого Рах6, Foxc1 и Pitx2 характеризуются более ограниченной зоной экспрессии в развивающемся глазу. Оба гена экспрессируются мезенхимой вокруг развивающегося глазного бокала (периокулярная мезенхима), в том числе клетками будущей роговицы. Их экспрессия снижается по ходу дифференцировки мезенхимы, и сохраняется в структурах развивающегося угла передней камеры. Foxc1 и Pitx2 оба экспрессируются в формирующейся радужке, но не экспрессируются в нейроэктодерме глазного бокала и хрусталикового пузыря.
г) Изучение функций генов на моделях животных. Каждый ген человека, мутации которого вызывают аномалии развития переднего сегмента глаза, имеет близко родственный и эквивалентный ген (ортологичный ген) в геноме мыши. У мышей, не имеющих функционирующих копий этих генов, развиваются те же аномалии переднего сегмента, которые вызывают развитие клинической картины у людей. Фенотипы мышей, гетерозиготных по мутации и имеющих лишь одну копию функционирующего гена, являются моделью клинических состояний с доминантным механизмом наследования.
Результаты исследований гомозиготных мутаций мышей, хотя они и редко встречаются в клинической практике, помогают выяснить важную роль ключевых генов. Существуют различия в генетической регуляции развития глаза у человека и у мыши, но мыши являются оптимальной моделью, исследование которой оказалось исключительно полезным для понимания природы аномалий развития переднего сегмента глаза и глаукомы.
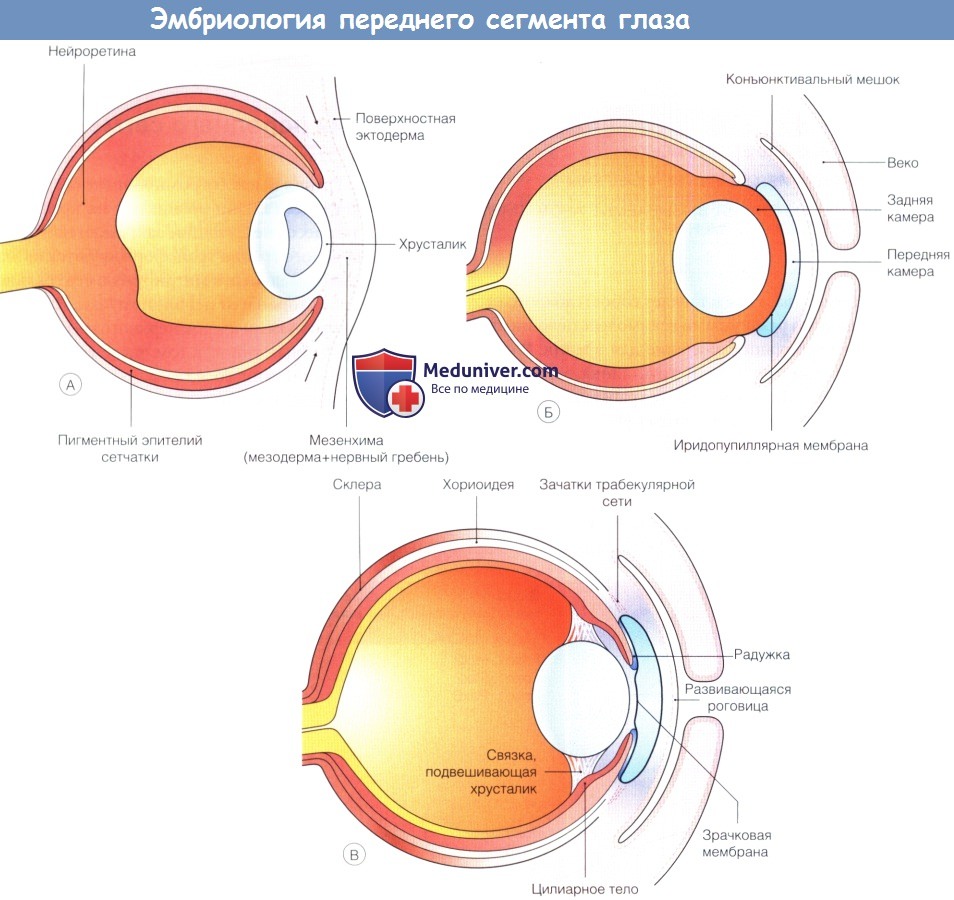
Раннее развитие переднего сегмента.
(А) Примерно пять недель развития эмбриона человека, хрусталиковый пузырек отделился от поверхностной эктодермы,
клетки нервного гребня мигрируют вокруг глазного бокала и между поверхностной эктодермой и развивающимся хрусталиком.
(Б) В течение седьмой недели из слоя мезенхимы развиваются выстланная эндотелием строма роговицы и передняя строма радужки.
Этот процесс дифференцировки идет одновременно с разделением двух слоев и формированием передней камеры между развивающимися роговицей и радужкой.
(В) Листок мезенхимы, перекидывающийся через будущий зрачок, сохраняется до седьмого месяца гестации.
Из краев глазного бокала образуется задний эпителий радужки и эпителий цилиарного тела.
д) Foxc1 и Pitx2 имеют большое значение для развития роговицы. Предполагается, что причиной развития аномалий радужки, появления тяжей ткани и аномалий линии Schwalbe при аномалиях развития переднего сегмента глаза является остановка развития, патологическая фиксация и сокращение эмбрионального эндотелиального слоя на участках радужки и в углу передней камеры.
Гетерозиготные мутации гена Foxcl у мышей вызывают аномалии развития глаз с очень вариабельной экспрессивностью; то же самое наблюдается в семьях, члены которых являются носителями одной и той же мутации, но имеют очень разные дефекты развития глаз. Дефекты развития глаз у гетерозигот постепенно прогрессируют и со временем корэктопия и периферические иридокорнеальные сращения становятся более заметными. У некоторых мышат выявлялось легкое помутнение роговицы, которое с высокой частотой у взрослых животных прогрессировало в выраженное помутнение с неоваскуляризацией, с сопутствующей катарактой.
При исследованиях на мышах также было выявлено, что генетическая среда влияет на пенетрантность дефектов развития глаза у гетерозигот и, вероятно, определяет различия клинической картины, наблюдаемые между семьями, представители которых являются носителями одних и тех же мутаций. Однако, поскольку высокая вариабельность отмечается и среди генетически идентичных мышей, различия в значительной степени должны возникать в результате особенностей развития, которые, в свою очередь, развиваются в результате случайных событий, связанных с уровнем ключевых молекул в критические моменты развития.
Асимметричные фенотипические признаки в разных глазах одного и того же пациента также, вероятно, являются результатом таких случайных событий.
У гомозиготных по мутантному Foxc1 мышей (которых называют врожденными гидроцефалами — congenital hydrocephalus, ch, или нацеленными на ген Мf1 — Мf1 gene-targeted) наблюдается более тяжелый дисгенез переднего сегмента. При гистологическом исследовании выявляется, что роговица не отделилась от хрусталика, в результате чего передняя камера полностью отсутствует. Наружный эпителий роговицы толще, чем в норме, строма дезорганизована. Эндотелий роговицы не дифференцирован, не сформированы плотные окклюзионные соединения между эндотелиоцитами, необходимые для осуществления нормальной физиологической барьерной функции. На фоне этого отсутствуют десцеметова мембрана и базальная пластинка, секретируемые эндотелиоцитами.
Аномалии роговицы обычно сопровождаются гипоплазией стромальной мезенхимы и пигментного слоя радужки, как правило, наблюдается микрофтальмия. Фенотип таких мышей указывает на важную роль Foxc1 в трансформации мезенхимальных клеток нервного гребня в эндотелиоциты.
Фенотип гомозиготных по мутантному гену Foxcl мышей неожиданно схож с фенотипом мышей—гомозиготных носителей мутации другого гена, определяющего появление аномалий развития переднего сегмента глаза у человека, гена Pitx2. У гомозиготных по мутантному Pitx2 мышей отмечаются смещенные зрачки неправильной формы, это состояние также выявляется у некоторых гетерозигот. У гомозигот передняя камера и эндотелий роговицы отсутствуют, эпителий роговицы утолщен (гиперклеточный), мезенхимальные клетки, лежащие между эпителием и глазным бокалом недифференцированы.
Pitx2, как оказалось, играет важную роль при дифференцировке мезенхимального и эпителиального компонентов роговицы, тканей, развивающихся из периокулярной мезенхимы — производной краниального нервного гребня и поверхностной эктодермы, соответственно. Экспрессия Pitx2, а не Foxc1, как сообщалось, выявлена в роговичной эктодерме (производной поверхностной эктодермы).
Отсутствие Pitx2 у мышей также препятствует развитию глазодвигательных мышц, вызывает уменьшение размеров глаза (микрофтальмия) и задержку закрытия эмбриональной глазной щели (колобома зрительного нерва). Эти аномалии не наблюдались у пациентов с мутацией PITX2.
У мышей, не имеющих Pitx2 также развиваются аномалии различных органов, являющихся важными зонами активности гена Pitx2. Pitx2 играет роль в формировании лево-правосторонней асимметрии, участвует в определении положения сердца, асимметрии легких и развитии гипофиза, зубов и краниофациальной зоны. У гетерозиготных животных выявляются лишь аномалии глаз и зубов, соответствующие аномалиям зубов пациентов с мутацией PITX2. Знание других органов, критически поражаемых при отсутствии Pitx2 полезно для трактовки других системных изменений, выявляемых у пациентов с дисгенезом переднего сегмента.
Недавно было продемонстрировано, что у человека фенотипы с тяжелыми сочетанными аномалиями развития переднего сегмента глаза могут определяться дигенным наследованием PITX2 и FOXC1.
е) Экспрессия Рах6 и других генов в хрусталике вызывает аномалии развития переднего сегмента глаза. В настоящее время точно установлено, что мутация или делеция гена РАХ6 и/или хромосомная перестройка, захватывающая ген РАХ6 на 11р13, является причиной большинства случаев аниридии.Дефекты РАХ6 также очень часто выявляются при мальформациях переднего сегмента.
Исследования гетерозиготных по мутантному Рах6 мышей помогают выяснить роль Рах6 в развитии аномалий переднего сегмента глаза, поскольку фенотип таких мышей сходен со связанными с мутацией РАХ6 аномалиями, наблюдаемыми у пациентов. У гетерозиготных по Рах6 мышей (мелкий глаз — small eye, Sey) отмечается уменьшение размеров глаз (микрофтальмия) и широкий спектр дефектов переднего сегмента глаза, в том числе гипоплазия роговицы, иридокорнеальные сращения и помутнение роговицы, неполное отделение хрусталика от роговицы (роговично-хрусталиковое сращение), васкуляризация роговицы и катаракта.
Аномалия Петерса, характеризующаяся роговично-хрусталиковыми и/или радужко-роговичными сращениями, является генетически гетерогенным состоянием. В части случаев аномалии Петерса имеются мутации РАХ6, и фенотип гетерозиготных мышей Sey напоминает фенотип при этой аномалии. Мутация FOXE3 также связана с аномалией Петерса, у гетерозиготных по мутантному Foxe3 мышей выявляется центральное помутнение роговицы и роговично-хрусталиковое сращение, как и при аномалии Петерса.
Хрусталик играет важную роль в индукции дифференциации переднего сегмента. Результаты исследования глаз гетерозигот по Рах6 указывают, что гаплонедостаточность Рах6 вызывает развитие первичных дефектов хрусталика, в свою очередь вызывающих вторичные сочетанные дефекты переднего сегмента и радужки. Рахб интенсивно экспрессируется передним эпителием хрусталика и, регулируя выработку хрусталиковых сигнальных молекул, может опосредованно воздействовать на мезенхимальные клетки развивающегося переднего сегмента — производные нервного гребня. Два других гена, нарушая индуктивные функции хрусталика, могут вызывать развитие аномалий переднего сегмента глаза.
Мутации MAF и FOXE3 вызывают аномалии развития переднего сегмента глаза и катаракты. У мышей гены Maf и Foxe3 первично экспрессируются развивающимся хрусталиком, но не мезенхимальными клетками развивающегося переднего сегмента — производными мезенхимальных клеток. В недавних работах было показано, что гомозиготные мутации FOXE3 вызывают первичную врожденную афакию.
Данные о различных функциях генов, вызывающих развитие аномалий переднего сегмента глаза, позволяют предложить модель, согласно которой Рахб, Maf и Foxe3 участвуют в выработке хрусталиком сигнальных факторов, играющих важную роль в управлении развитием переднего сегмента. Pitx2, Foxc1 и Lmx1b имеют большое значение для дифференцировки мезенхимальной ткани — производной нервного гребня, регулируемой секретируемыми хрусталиком факторами. Без этих экспрессируемых мезенхимой генов не происходит разделения роговицы и хрусталика, формирования передней камеры и дифференциации дренажной системы угла передней камеры, либо в зависимости от дозы гена эти процессы происходят не полностью.
ж) Предположения об этиологии глаукомы при аномалиях развития на основании изучения модели мыши с аномалиями развития переднего сегмента глаза. Взаимосвязи мутаций генов, структурных аномалий угла передней камеры и высокой частоты глаукомы при аномалиях развития переднего сегмента глаза до конца не выяснены. По результатам гистологического исследования угла передней камеры таких пациентов выявлено нарушение развития межтрабекулярных пространств и шлеммова канала. Анализ мышиной модели патологии человека убедительно показывает, что моногенные мутации вызывают аномалии ткани трабекулярной сети и обструкцию оттока водянистой влаги.
У гомозиготных по Foxc1 мышей при гистологическом исследовании радужко-роговичного угла выявляются такие аномалии, как малый размер или отсутствие шлеммова канала, гипоплазия или отсутствие трабекулярной сети и гипоплазия цилиарного тела с короткими и тонкими цилиарными отростками. Развитие угла передней камеры также изучалось у мышей — гетерозигот по Рах6. Мезенхимальные клетки угла передней камеры, в норме экспрессирующие Рах6 и дифференцирующиеся в прилегающую к шлеммову каналу трабекулярную сеть, остаются недифференцированными, что свидетельствует о значении Рах6 для дифференциации утла передней камеры. В дополнение к нарушениям развития, приводящим к дефектам переднего сегмента глаза и глаукоме, появилась новая концепция первичной врожденной глаукомы и генетических механизмов, вызывающих эти близкие друг другу заболевания.
У мышей, у которых отсутствует Сур1b1, имеются те же фокальные аномалии развития угла передней камеры, что описаны у пациентов с первичной врожденной глаукомой и мутацией CYP1B1, а именно: узкий шлеммов канал или его отсутствие, базальная пластинка (напоминающая десцеметову мембрану), простирающаяся от роговицы за трабекулярную сеть, сращения радужки, трабекулярной сети и периферических отделов роговицы (синехии).