Глазодвигательные нарушения у младенцев и маленьких детей бывают врожденными или приобретенными, связанными с нарушениями раннего развития зрительной системы, а также могут являться проявлениями фоновых неврологических или нервно-мышечных заболеваний или патологии глазницы. Аномальные движения глаз у внешне здорового ребенка никогда не следует относить на счет врожденного или доброкачественного состояния, не проведя тщательного обследования, включающего в себя сбор анамнеза, клинический осмотр, лучевые исследования, лабораторные анализы и электрофизиологическое обследование зрительной сенсорной системы. Анализ движений глаз может указать на тип глазодвигательного расстройства и на фоновое заболевание глаз или неврологическую патологию.
а) Нейроинтегратор. Нейроинтегратор — это технический термин, которым обозначают глазодвигательную «функцию», выполняемую группами клеток мозжечка (клочка — flocculus и околоклочковой дольки — paraflocculus), предлежащего подъязычного ядра (prepositus hypoglossus) и медиального вестибулярного ядра. Нейроинтегратор необходим для всех содружественных движений глаз. Чтобы двигать глаза с постоянной скоростью или удерживать их в эксцентрической позиции взора, два нервных сигнала должны противодействовать эластичной тяге, стремящейся вернуть глаза в положение «покоя». Эти сигналы определяют: один — необходимую скорость (фазовый компонент), а другой — тонический компонент, уравновешивающий эластическое противодействие.
Тончайшие изменения тонического компонента регулируются премоторным нервным сигналом, который математически «интегрирует» сигнал скорости — этот механизм и был назван нейроинтегратором. Роль нейроинтегратора в развитии саккад и поддержании эксцентрических положений взора реализуется при подавлении ингибиторных взрывных нейронов (inhibitory burst neurons, IBN) и быстрых разрядах возбуждающих взрывных нейронов (excitatory burst neurons, EBN), в результате чего возникает интенсивный фазовый сигнал, передающийся по соответствующим черепным нервам на конкретную сопряженную пару глазодвигательных мышц. «Взрывной» сигнал также передается на нейроинтегратор, который «интегрирует» этот взрывной сигнал (подсчитывается количество пиков разрядов) и генерирует нервный сигнал (также передаваемый по черепным нервам), регулирующий фиксацию глаза в новой позиции (тонический разряд).
Нейроинтегратор не является совершенной системой, и «тонический» сигнал медленно ослабевает или «рассеивается» со временем. В фотопических условиях это ослабление в норме не заметно из-за зрительной обратной связи: система плавного слежения/фиксации способствует стабилизации положения глаза. При рождении нейроинтегратор еще не сформирован, но примерно к месячному возрасту он уже функционирует.
б) Анатомия и физиология системы саккад. Саккады представляют собой быстрые движения глаз, во время которых центральная ямка целенаправленно поворачивается в сторону конкретного объекта; они могут возникать произвольно (произвольные саккады), рефлекторно, или быть частью быстрофазовых движений при нистагме. Произвольные саккады могут возникать как проявление предварительных команд (например при взгляде вправо), саккады по памяти (memory-guided saccades) и антисаккады. Непроизвольные саккады включают в себя быстрофазовые движения при нистагме, спонтанные саккады и рефлекторные саккады. Путь саккад проходит через переднее бедро внутренней капсулы, через промежуточный мозг, а затем разделяется на дорсальный и вентральный пути: дорсальная порция идет в верхние бугры четверохолмия, а вентральная порция (содержащая глазодвигательные пути горизонтальных и вертикальных движений глаз) в мост и средний мозг. Верхний бугор четверохолмия функционирует как важный переключатель (реле) некоторых из этих проекций.
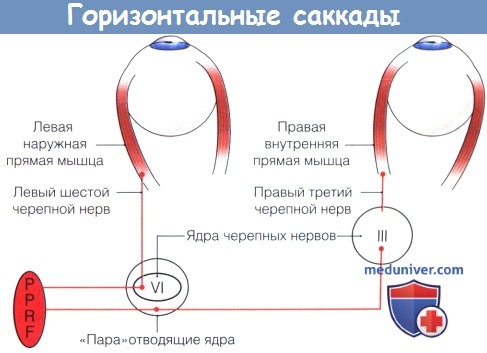
Ростральное интерстициальное ядро медиального продольного пучка (rostral interstitial nucleus of the medial longitudinal fasciculus — riMLF) и парамедианная ретикулярная формация моста (pontine paramedian reticular formation — PPRF) ствола головного мозга определяют скорость саккад: непосредственно перед движением глаза они генерируют «пульсирующие нервные импульсы», идущие по III, IV и VI черепным нервам. Горизонтальные саккады генерируются возбуждающими взрывными нейронами парамедианной ретикулярной формации моста, обнаруженными у вентральной и латеральной стороны медиального продольного пучка моста, и ингибиторными взрывными нейронами дорсального парагигантоклеточного ядра, лежащего каудальнее ядра отводящего нерва в ростральной дорсомедиальной части продолговатого мозга. Вертикальные и ротаторные компоненты саккад определяются импульсами, генерируемыми возбуждающими и ингибиторными взрывными нейронами рострального интерстициального ядра медиального продольного пучка, локализующимися в среднем мозге.
Вслед за саккадой происходит «переключение иннервации», в результате которого на глазодвигательные мотонейроны воздействует тоническая иннервация более высокого уровня и глаз удерживается в новом положении, вопреки эластическим силам, стремящимся вернуть глаз в анатомически «нейтральное» положение в глазнице. После переключения иннервации горизонтальные саккады контролируются N1 (nucleus interpositus мозжечка), наиболее важное влияние оказывают предлежащее ядро — комплекс медиального вестибулярного ядра. По окончании вертикальных и ротаторных саккад глаза удерживаются в устойчивом положении благодаря переключению иннервации на промежуточное ядро Cajal среднего мозга. Другие взрывные нейроны, называемые замедленными взрывными нейронами (LLBNs, long-lead burst neurons), генерируют импульс за 40 мс до саккады, тогда как возбуждающие взрывные нейроны генерируют импульс за 12 мс до саккады. Некоторые замедленные взрывные нейроны локализуются в среднем мозге, они получают проекции от верхнего бугра четверохолмия и отдают проекции в возбуждающие взрывные нейроны моста, ингибиторные взрывные нейроны продолговатого мозга и OPNs (omnipause neurons — «универсальные паузовые нейроны»).
Другие замедленные взрывные нейроны (LLBNs) лежат в ретикулярном ядре покрышки моста и отдают проекции в основном в мозжечок, но также и в парамедианную ретикулярную формацию моста. Представляется, что замедленные взрывные нейроны (LLBNs), получающие входящую им-пульсацию от верхних бугров четверохолмия, могут играть ключевую роль в трансформации пространственной кодировки команд во временную кодировку, тогда как другие замедленные взрывные нейроны (LLBNs) могут синхронизировать начало и конец саккад.
Система саккад полностью развивается примерно к окончанию первого года жизни; для фиксации объекта младенцы выполняют множество гипометрических саккад. У младенцев младше трех месяцев, при неподвижной головке, эти гипометрические саккады можно наблюдать клинически, особенно при больших саккадах. В течение первых семи месяцев жизни наблюдается прогресс в сторону «нормометрии». У здоровых взрослых и детей младше одного года, саккады обычно гипометрические, их протяженность составляет около 90-100% дистанции до цели, после чего наблюдаются вторичные саккады — нормометрия. При оптокинетическом нистагме и вертикальном нистагме быстрые фазы у детей возникают реже, чем у взрослых.
Схема путей ствола головного мозга, координирующих горизонтальные саккады.
Парамедианная ретикулярная формация моста (PPRF) получает входящие импульсы от ипсилатерапьных корковых центров и верхнего бугра четверохолмия и стимулирует две группы нейронов отводящего ядра:
(1) их аксоны идут к подгруппе нейронов, иннервирующих ипсилатеральную наружную прямую мышцу, и
(2) аксоны которых объединяются с медиальным продольным пучком, после чего активируют подгруппу нейронов, иннервирующих внутреннюю прямую мышцу контралатерального третьего черепного нерва.
PPRF, paramedian pontine reticular formation — парамедианная ретикулярная формация моста, VI—ядра шестого черепного нерва, III—ядра третьего черепного нерва.
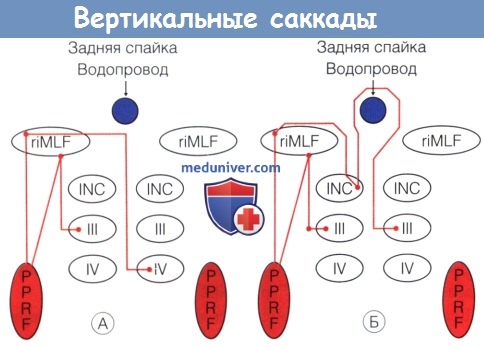
Схема путей ствола головного мозга, координирующих саккады, направленные вниз (А) и вверх (Б).
(А) Парамедианная ретикулярная формация моста активирует нейроны рострального интерстициального ядра медиального продольного пучка,
от которого идут волокна в каудальном направлении к синапсам нейронов подгруппы нижней прямой мышцы ипсилатерапьного третьего черепного нерва и ядра контралатерального блокового нерва.
Не показанные на схеме волокна от контралатеральной парамедианной ретикулярной формации моста одновременно несут согласовывающие сигналы.
(Б) Парамедианная ретикулярная формация моста активирует нейроны рострального интерстициального ядра медиального продольного пучка,
волокна от которых идут через заднюю спайку к подгруппе нейронов верхней прямой мышцы контралатерального третьего черепного нерва и к подгруппе нейронов нижней косой мышцы ипсилатерапьного третьего черепного нерва.
Не показанные на схеме волокна от контралатеральной парамедианной ретикулярной формации моста одновременно несут согласующие сигналы.
riMLF, rostral interstitial nucleus of the medial longitudinal fasiculus — ростральное интерстициальное ядро медиального продольного пучка, INC, interstitial nucleus of Cajal—промежуточное ядро Cajal,
III — ядро третьего черепного нерва, IV — ядро четвертого черепного нерва, PPRF, paramedian pontine reticular formation—парамедианная ретикулярная формация моста.
д) Анатоми и физиология системы плавного слежения глаз. Функция плавного слежения (smooth pursuit — SP) заключается в сохранении фиксации движущегося объекта, для чего необходимы движения обоих глаз и головы. Для осуществления плавного слежения требуется способность к прогнозированию, позволяющая компенсировать инерционность визомоторной системы, и возможность подавлять вестибулоокулярный рефлекс. Необходимость подавления вестибулокоулярного рефлекса обусловлена тем, что при движении головы возникающий вестибулоокулярный рефлекс вызывает поворот глаз в противоположную сторону. Плавное слежение и подавление вестибулоокулярного рефлекса, вероятно, является одной глазодвигательной функцией. Фронтальные и экстрастриарные зоны зрительной коры передают информацию о движении и объекта, и глаз в дорсолатеральные ядра моста, оттуда идут связи в клочок и околоклочковую область (paraflocculus, flocculus) дорсальной части червя, а оттуда через вестибулярные ядра и ядру шатра — к глазодвигательным ядрам III, IV и VI черепных нервов. Односторонние поражения коры и мозжечка вызывают нарушения ипсилатеральных плавных следящих движений. Клинические проявления стволовых поражений не столь специфичны.
Плавные следящие движения имеются уже в первую неделю жизни, но у младенцев они еще незрелы. Горизонтальные (плавные движения) взора, возможно, развиваются раньше вертикальных. Способность к плавному слежению развивается с возрастом, и в пять месяцев плавные следящие движения уже хорошо заметны. В каком возрасте способность к слежению созревает до взрослого уровня развития, неизвестно. Представляется, что это происходит не ранее шести месяцев жизни, хотя возможно, что ее разви тие завершается только в старшем подростковом возрасте.
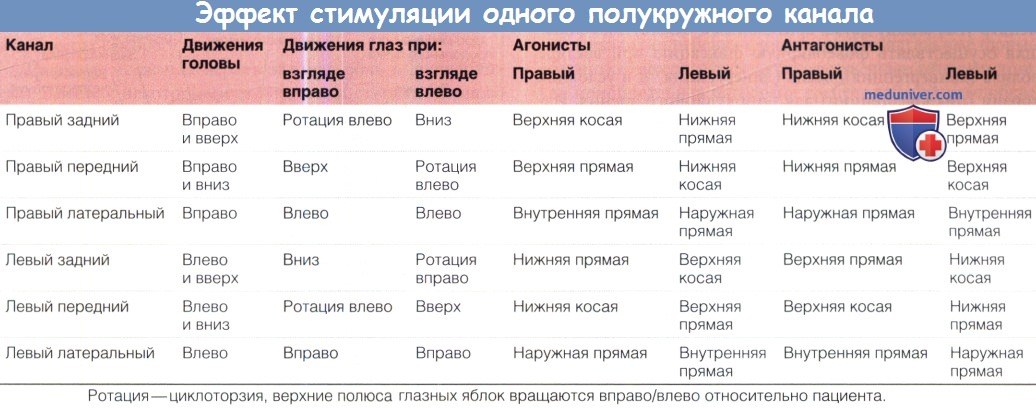
е) Система вестибулоокулярных реакций. Вестибулярный аппарат запускает глазодвигательные рефлексы, которые позволяют при движениях головы устойчиво удерживать изображение на сетчатке. Глаза поворачиваются в направлении, противоположном направлению поворота головы таким образом, что они не меняют своего положения в пространстве. Основные связи образуют трехнейронную дугу — вестибулярный ганглий, вестибулярные ядра и глазодвигательные ядра. Прямые нейрональные пути включают в себя тормозные и возбуждающие компоненты. Каждый полукруглый канал оказывает воздействие на пару глазодвигательных мышц, поворачивающих глаз в плоскости этого канала. Подробно описана анатомия вестибулярных ядер; они получают проекции от 14000-18000 аксонов вестибулярного нерва.
Существует четыре основных вестибулярных ядра и добавочные подгруппы, в том числе интерстициальное ядро, клетки которого распределены между вестибулярными корешками при их вхождении в ствол головного мозга, и у-группу вблизи верхней ножки мозжечка. Медиальное вестибулярное ядро имеет самый большой объем и является наиболее длинным из вестибулярных ядер. Латеральное вестибулярное ядро также отдает проекции в спинной мозг, в основном через ипсилатеральный латеральный вестибулоспинальный тракт, но также через контралатеральный медиальный вестибулоспинальный тракт. Наиболее ростральная часть нижнего вестибулярного ядра также отдает проекции в глазодвигательные ядра.
Первичные вестибулярные афферентные пути входят в продолговатый мозг на уровне латерального вестибулярного ядра. Почти все они разделяются надвое, нисходящая ветвь идет в медиальное вестибулярное ядро и нижнее вестибулярное ядро, восходящая ветвь — в верхнее вестибулярное ядро; заканчиваются вестибулярные афферентные пути в мозжечке, большая часть — в передней части червя, в его узелке (nodulus) и втулочке (uvula). Все каналы и отолиты отдают проекции к границам вентромедиальной части латерального вестибулярного ядра, медиальной части медиального вестибулярного ядра и дорсомедиальной части нижнего вестибулярного ядра. Импульсация от всех каналов также сходится в маленьком участке вентромедиальной части верхнего вестибулярного ядра. Маточка отдает проекции в ростральную часть медиального вестибулярного ядра, а мешочек — в у-группу.
И при горизонтальных, и при вертикальных вестибулоокулярных рефлексах множество нейронов вестибулярных ядер получают первичные афферентные вестибулярные импульсы и кодируют скорость поворота головы, положение глаз и различные параметры сигнала следящих движений и саккад. Нейроны вестибулярных ядер отдают проекции не только в мотонейроны; также от них идут коллатерали к предлежащему подъязычному ядру (nucleus prepositus hypoglossi), ядру Roller, и группам клеток парамедианного тракта.
При исследовании вестибулоокулярных рефлексов у младенца нередко можно наблюдать немного усиленный вестибулоокулярный ответ, постепенно ослабевающий в дошкольном возрасте, и короткий период затухания вестибулоокулярной реакции. У недоношенных младенцев и у некоторых здоровых доношенных младенцев вращение вызывает тоническую девиацию или «замыкание». При выполнении теста «кукольной головы» или вращении на кресле Вагапу глаза отклоняются в направлении, противоположном направлению вращения. При вращении ребенка на вытянутых руках его глаза отклоняются в направлении вращения. «Корректирующая» быстрая фаза развивается приблизительно на 45 неделе постконцептуального возраста. Подобное «замыкание» у младенца с задержкой зрительного развития может сохраняться в течение и более длительного периода. Обычно после окончания вращения наблюдается всего несколько колебаний постротаторного нистагма; если наблюдается большее количество колебаний нистагма, следует подозревать тяжелые зрительные нарушения или аномалию путей плавного слежения. На результаты этого теста могут влиять настроение и утомление ребенка.
ж) Анатомия и физиология системы вергенции глаз. Вергенцией называется одновременное движение глаз в противоположных направлениях, за счет которого изображения объектов попадают на корреспондирующие точки сетчатки. Другими словами, вергенцией называется способность изменять угол между двумя зрительными осями, позволяющая осуществлять фовеолярную фиксацию при взгляде вблизь (конвергенция), вдаль (дивергенция) и в условиях торзии (цикловергенции) для бинокулярного зрения. Известны три основных стимула, вызывающих вергенцию:
(1) диспарантность сетчатки вызывает фузионную вергенцию;
(2) размытость изображения на сетчатке вызывает аккомодационную вергенцию; и
(3) движение вызывает и диспарантность, и аккомодационную вергенцию.
Нейронный субстрат вергенции локализуется в ретикулярной формации среднего мозга, с дорсолатеральной стороны от глазодвигательного ядра; импульсация этих нейронов определяет угол вергенции (клетки тонуса вергенции), скорость (взрывные клетки вергенции) или и угол, и скорость (взрывные-тонические клетки вергенции). Хотя большинство из этих нейронов также посылают импульсы и при аккомодации, некоторые связаны преимущественно с вергенцией. Как и для верзивных движений глаз, необходима пространственно-скоростная интеграция вергентных сигналов: важную роль в этих интеграционных процессах играет ретикулярное ядро покрышки моста (nucleus reticularis tegmcnti ponds — NRTP). В ретикулярном ядре покрышки моста клетки, участвующие в реакциях на взгляд вблизь, лежат отдельно от клеток, осуществляющих реакции при взгляде вдаль. Поражение ретикулярного ядра покрышки моста приводит к неспособности устойчиво удерживать угол вергенции. Ретикулярное ядро покрышки моста связано реципрокными связями с мозжечком (межпозиционное ядро — nucleus interpositus) и получает нисходящие проекции от нескольких корковых и подкорковых структур.
Часто кажется, что глаза новорожденных, особенно недоношенных, дивергируют, так как до 2-3 месячного возраста произвольная конвергенция слаба. В возрасте от трех до шести месяцев у 75% недоношенных и 97% доношенных младенцев девиации отсутствуют. Фузия полностью развивается только к шестимесячному возрасту. Аккомодационную вергенцию можно обнаружить в двухмесячном возрасте, а диспарантную вергенцию (disparity driven vergence) — в возрасте около четырех месяцев, т.е. в период развития стереопсиса и фузии.
з) Анатомия и физиология оптокинетического нистагма. Оптокинетическая система отвечает за содружественное медленное слежение за движущимися большими участками поля зрения. Оптокинетический нистагм (optokinetic nystagmus — OKN) представляет собой рефлекторный содружественный физиологический нистагм, при котором за медленной фазой реакции слежения за движущейся окружающей средой (оптокинез — optokinesis) следует быстрая фаза — корректирующая саккада. Оптокинетический нистагм возникает в естественных условиях при движении головы и глаз и в искусственных условиях при взгляде из окна движущегося транспортного средства. Во взаимодействии с вестибулярной системой оптокинетическая система обеспечивает неподвижность изображения на сетчатке во время непрерывного движения головы или окружающего мира, или и того, и другого.
Нейронный субстрат оптокинеза состоит из ядра зрительного тракта и добавочных зрительных путей. Оптокинез может вызываться движением во всем поле зрения (естественные движения головы или глаз) или движением в небольшом участке поля зрения (центральной ямке) (вращающийся барабан или движущаяся лента). И система плавного слежения, и оптокинетическая система участвуют в стабилизации изображений стационарных объектов при поворотах головы.
У приматов оптокинетический нистагм состоит из двух компонентов, каждый из которых имеет отдельный нервный путь, хотя и параллельный другому.
Отсроченный (непрямой, медленный) оптокинетический нистагм (delayed OKN — OKNd) развивается медленно (десятки секунд) и переходит в оптокинетический постнистагм (optokinetic after-nystagmus, OKAN), представляющий собой постепенное исчезновение нистагма после выключения света. Отсроченный оптокинетический нистагм тесно связан с вестибулоокулярным рефлексом и управляется визомоторными сигналами зрительной коры, идущими через ядро зрительного тракта претектальной области и вестибулярные ядра. Ранний (прямой, быстрый) оптокинетический нистагм (early OKN — OKNe) характеризуется быстрым началом (< 1 секунды) и не переходит в оптокинетический постнистагм; в темноте он быстро прекращается. Путь раннего оптокинетического нистагма аналогичен пути плавного слежения, он проходит через кортикопонтоцеребеллярный путь, участие претектальной области в процессах раннего оптокинетического нистагма/плавного слежения сомнительно, хотя она может принимать участие в процессах саморегуляции.
Наконец, мозжечок играет важную роль в движениях глаз. Вместе с некоторыми структурами ствола головного мозга, включая предлежащее ядро и медиальное вестибулярное ядро, он, как оказывается, посредством математической интеграции конвертирует скоростные сигналы в сигналы положения при всех содружественных движениях глаз.
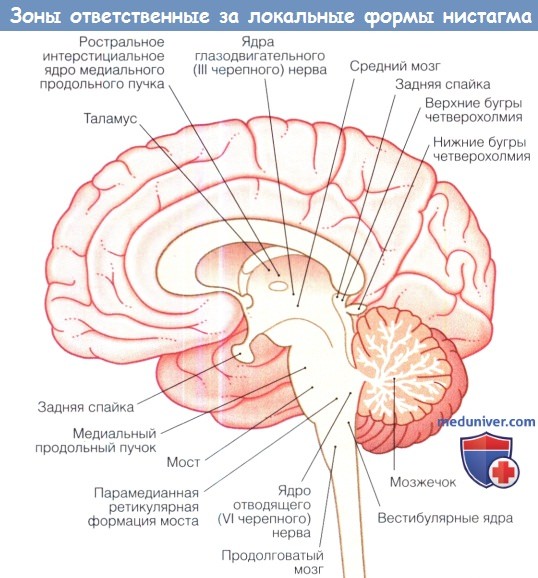
Срединный сагиттальный разрез головного мозга человека.
Зоны головного мозга, ответственные за развитие «локальных» форм приобретенного нистагма.