• Селективная проводимость и воротный механизм (гейтинг) в Cl--каналах структурно сопряжены
• В К+- и Cl--каналах используются различные механизмы гейтинга и селективности
Хлоридные каналы (Cl-) относятся к большой группе анионных каналов. Как и другие ионные каналы, канальные белки хлоридных каналов образуют поры в биологических мембранах. Cl--каналы обеспечивают транспорт отрицательно заряженных ионов Cl- в направлении их электрохимического градиента.
В экспериментах in vitro Cl--каналы функционируют как неселективные анионные каналы, которые иногда способны транспортировать другие анионы даже лучше, чем Cl-. Однако Cl- относится к наиболее распространенным анионам клетки, и поэтому in vivo по таким каналам, главным образом, осуществляется транспорт Cl- анионов.
Хлоридные каналы находятся в плазматической мембране и в мембранах внутриклеточных органелл. К числу их важнейших функций относится регуляция клеточного объема, ионного гомеостаза и трансэпителиального транспорта ионов. Транспорт Cl- через плазматическую мембрану играет важную роль в регуляции возбудимости мышц и нейронов. При закислении внутриклеточных компартментов, например эндосом, транспорт в них Cl- по соответствующим каналам нейтрализует положительный заряд протонов, переносимых Н+-АТФазами.
Гены, кодирующие белки Cl--каналов, относятся к трем различным семействам. Первая группа — гены семейства CLC, включает несколько генов, кодирующих белки плазматической мембраны, а также мембран внутриклеточных компартментов. CLC-каналы присутствуют во всех организмах — от бактерий до человека и, в свою очередь, подразделяются на подгруппы по принципу гомологии. Ко второй группе относятся АВС-переносчики, функционирующие как ионные каналы.
Единственный представитель этой группы — регулятор трансмембранного переноса при кистозном фиброзе. К третьей группе принадлежат рецепторы у-аминомасляной кислоты, находящиеся под контролем лигандов, и глициновые рецепторы. Все они являются представителями группы Cl--каналов, обладающих специализированными функциями в ЦНС.
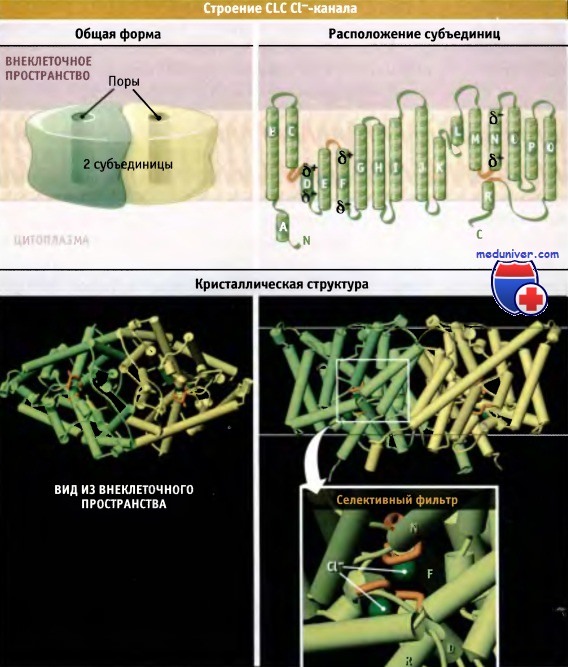
Схематическое строение бактериального транспортного CLC Cl--комплекса и его структура по данным рентгенокристаллографического исследования.
Cl--транспортные белки из семейства CLC образуют гомодимеры, и каждая субъединица образует отдельную пору.
Части белковой субъединицы, образующие селективный фильтр, выделены оранжевым цветом.
Модель кристаллической структуры построена на основании данных Protein Data Bank file 1KPK.
Вид из внеклеточного пространства по масштабу меньше, чем вид сбоку. Показано предполагаемое положение мембраны.
CLC Cl--каналы представляют собой гомодимеры, каждая субъединица которых образует отдельную проводящую пору. Рентгеноструктурный анализ CLC Cl--каналов клеток эукариот не проводился, и мы не располагаем данными об их атомарной структуре. Однако охарактеризована кристаллическая структура соответствующих бактериальных белков.
Эти белки функционируют как переносчики, транспортирующие Cl--ионы в обмен на протоны, а не в качестве хлоридных каналов, как это предполагалось ранее. Однако результаты секвенирования бактериальных переносчиков демонстрируют их сходство с CLC Cl--каналами эукариот. Действительно, CLC-переносчики бактерий представляют собой также гомодимеры. Каждая субъединица состоит из 18 а-спиралей, собранных в сложный трансмембранный комплекс.
N-терминальная половина (спирали A-I) одной субъединицы близка по структуре к С-терминальной половине (спирали J-R). Две половины субъединицы обладают противоположной (антипараллельной) мембранной ориентацией, напоминающей структуру аквапориновых каналов.
Важнейшим свойством всех CLC Cl--каналов является внутренняя сопряженность ионной проводимости и воротного механизма. Это следует из структуры селективного фильтра CLC прокариотических клеток. В центре белковой структуры, в самой узкой части поры, расположен селективный фильтр, а устье поры имеет форму часового стекла. Селективность канала, которая обусловливает преимущественный транспорт по нему анионов, связана с двумя особенностями его строения.
Первая состоит в том, что положительно заряженные N-концевые группы четырех а-спиральных сегментов (D, F, N и R) обращены к центральной плоскости мембраны. Вторая особенность касается аминокислотных остатков, которые облицовывают пору. Они образуют водородные связи с Cl--ионами. Такое строение поры создает благоприятное электростатическое окружение для ионов Cl- за счет стабилизации их в селективном фильтре и отталкивания положительно заряженных катионов.
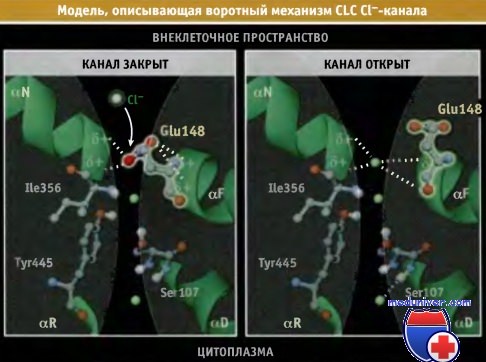
Предполагается, что роль ворот выполняет боковая цепь остатка глутаминовой кислоты, расположенная на одной стороне селективного фильтра. Механизм работы ворот становится понятным при анализе кристаллической структуры бактериальных CLC-переносчиков дикого типа и их мутантных форм, у которых глутамат заменен остатком другой аминокислоты.
В закрытом канале боковая цепь остатка глутамата занимает сайт связывания Cl-, имитируя анион. При открытии боковая цепь отклоняется в сторону, тем самым освобождая канал, и ее место занимает хлорид-ион. Остаток глутаминовой кислоты присутствует почти во всех CLC-каналах. Поэтому можно предполагать, что селективный фильтр CLC-переносчика у бактерий имеет такое же строение, как и CLC Cl--канал клеток эукариот
Селективный фильтр Cl--канала содержит три сайта связывания хлорид-ионов.
В закрытом канале боковая цепь глутаминовой кислоты (Glul48) занимает сайт связывания иона Cl-, который ближе всего расположен к устью поры.
При открытии канала боковая цепь отклоняется и выходит из канала, а в него поступает хлорид-ион.
Показаны остатки аминокислот, участвующие в позиционировании двух других хлорид-ионов.
Модели построены на основании данных по кристаллической структуре CLC-канала Е. coli, Protein Data Bank file 10TS.
Что вызывает вращение остатка глутамата, который играет роль ворот? Различные хлоридные каналы контролируются лигандами, электрическим потенциалом или изменениями концентрации Са2+ в клетке. Они также могут активироваться за счет градиента ионов Cl-. Модель, изображенная на рисунке ниже, предполагает, что при превышении определенной внеклеточной концентрации ионов Cl- они вытесняют глутамат из сайтов связывания анионов и начинают транспортироваться через канал. Таким образом, хлорид-ионы и отрицательно заряженные карбоксильные группы боковой цепи глутамата взаимно отталкиваются и конкурируют между собой за связывание с частичными положительными зарядами на а-цепи N.
Это обеспечивает быстрый транспорт анионов через канал и дает возможность регулировать воротный механизм непосредственно с помощью хлорид-ионов. Поскольку изменения заряда на мембране могут привести к усилению электрохимического потенциала трансмембранных градиентов Cl-, эта модель также объясняет потенциал-зависимое открытие большинства CLC Cl--каналов. Эти каналы не имеют заряженных трансмембранных доменов, выполняющих роль сенсоров заряда, подобно тем, которые находятся в потенциал-зависимых катионных каналах.
Cl-- и К+-каналы обладают принципиально различными механизмами, которые обеспечивают транспорт и селективность отбора ионов. Во-первых, в CLC Cl--каналах селективный фильтр и ворота представляют собой одну структурную единицу. Напротив, в К+-каналах селективный фильтр и ворота образованы двумя раздельными структурами, находящимися на наружной и внутренней сторонах канала соответственно. Такое разделение двух компонентов канала дает возможность домену, связывающему лиганд, или сенсору заряда открывать и закрывать поры за счет конформационных изменений.
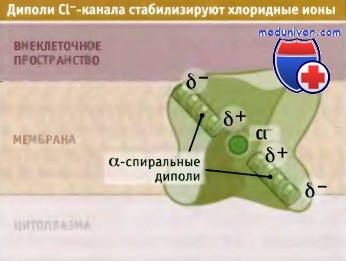
Структура селективного фильтра при этом не меняется, и его способность различать катионы, лишь незначительно отличающиеся по размеру, сохраняется. Во-вторых, воротный механизм хлоридного канала предполагает более ограниченное движение (вращение боковой цепи глутамата), чем то, которое происходит при существенных конформационных изменениях в К+-канале. В-третьих, необходимое электрическое окружение для транспортируемых ионов Cl- и К+ создается частичными положительными или отрицательными зарядами соответственно. Как показано на рисунке ниже для того чтобы направить частичный положительный заряд диполя N-терминальной спирали к селективному фильтру, CLC Cl--каналы имеют антипараллельную структуру. Напротив, для структуры К+-каналов характерна параллельная направленность, при которой частичный отрицательный заряд диполя С-терминальной спирали направлен на заполненную водой полость, представляющую собой часть их селективного фильтра.
Две половины субъединицы CLC Cl--канала находятся в антипараллельной ориентации таким образом,
что положительнозаряженные участки диполей определенных а-спиралей обращены в сторону селективного фильтра и влияют на связывание аниона Cl- с обеих сторон мембраны.
Показана одна субъединица.
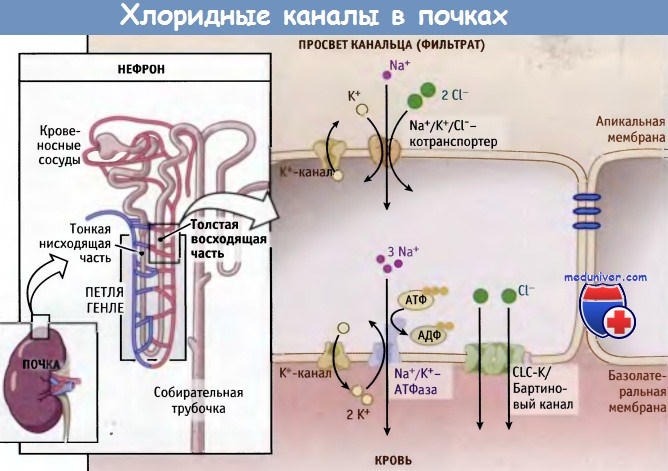
Роль Cl--каналов в трансэпителиальном транспорте ионов можно проиллюстрировать на примере функционирования CLC К-каналов почек. Для экспрессии этих каналов на базолатеральной мембране толстой восходящей части петли Генле необходима ассоциация с b-субъединицей бартина. Через восходящую часть петли Генле реабсорбируется 25% ионов (NaCl и К+), однако она непроницаема для воды. Для реабсорбции NaCl особенно необходимы CLC К-каналы, и они также косвенным образом поддерживают необходимую концентрацию К+ в плазме крови. Это особенно важно, поскольку нервная система и сердце очень чувствительны к изменениям содержания ионов К+.
Почки фильтруют из крови конечные продукты обмена и ионы. В ходе этого процесса в фильтрат плазмы, который является первичной мочой, переходят анионы и катионы. В дальнейшем они должны реабсорбироваться с тем, чтобы их концентрация в крови поддерживалась на необходимом уровне. Ионы Na+ и Cl- реабсорбируются из фильтрата в дистальном прямом канальце — эпителиальном тубулярном сегменте толстой восходящей части петли Генле. Реабсорбция хлорид-ионов из фильтрата происходит в апикальной мембране посредством Na+/К+/Cl--котранспортеров. Транспорт осуществляется за счет действия базолатеральной Na+/К+-АТФазы, которая создает трансмембранный градиент ионов Na+, выкачивая их из клетки. Низкое внутриклеточное содержание ионов Na+ создает благоприятный электрохимический градиент для протекания транспортных процессов, сопряженных с Na+.
Реабсорбция хлорид-ионов происходит при участии апикальных Na+/К+/Cl--котранспортеров и приводит к увеличению концентрации в клетке ионов Cl-. Таким образом, их концентрация становится выше, чем во внеклеточной среде и в просвете канальца, и они начинают выходить из клетки в направлении градиента. Хлорид-ионы выходят в кровь через ClC-K-Cl--каналы базолатеральной мембраны. Ионы К+, которые проникли в клетку через Na+/К+/Cl--котранспортеры и базолатеральную Na+/К+-АТФазу, выходят из клетки через К+-каналы в апикальной и базолатеральной мембране. Без ClC-K-Cl- каналов поток ионов через Na+/К+/Cl--котранспортеры был бы недостаточным и привел бы к снижению абсорбции NaCl и ионов К+.
В противоположность большинству клеток млекопитающих, для которых значение мембранного потенциала покоя определяется проводимостью ионов К+, в скелетных мышцах примерно 80% значения мембранного потенциала покоя обеспечивается ClC-1 изоформой хлоридного канала. Важная роль различных CLC Cl--каналов и других белков транспорта ионов была установлена в экспериментах по сайт-специфическому мутагенезу при развитии болезней человека и на нокаутных мышах, у которых отсутствует экспрессия специфических изоформ Cl--канала. Например, мутации, приводящие к инактивации ClC-1-канала, вызывают различные формы длительного сокращения мышц, получивших название миотонии.
При развитии миотонии аномальная раздражимость вызывает напряженность скелетных мышц и невозможность их расслабления. Мутации в генах, кодирующих синтез белков ClC-К-хлоридных каналов или субъединиц бартина, которые экспрессируются в клетках почек и некоторых отделов преддверно-улиткового органа, приводят к развитию аутосомно-рецессивного наследственного заболевания, называемого синдром Бартера. При этом заболевании не достигается достаточно полная реабсорбция ионов СГ из мочи, что приводит к снижению уровня К+ в плазме, метаболическому алкалозу, развитию компенсаторного гиперальдостеронизма (при котором организм старается преодолеть последствия снижения объема плазмы и низкого кровяного давления), а также к сенсорно-невральной глухоте.
При эндоцитозе, а также при эндосомальном закислении функционирует ClC-5-изоформа внутриклеточного хлоридного канала. Мутации в соответствующем гене приводят к болезни Дента, которая характеризуется избыточным выведением Са2+ и белка с мочой и образованием почечных камней. Для обеспечения электронейтральности среды при кислотной секреции мембраны остеокластов содержат ClC-7 Cl--каналы, которые функционируют совместно с Н+-АТФазами, закисляющими среду. Мутации в изоформе ClC-7 хлоридного канала приводят к нарушению резорбции кости, сопровождающемуся сжатием костномозговой полости и ее деформацией, называемой остеопетроз.
Трансэпителиальный транспорт NaCl в почках.
Эпителиальные клетки специализированных участков нефрона (толстая восходящая часть петли Генле)
реабсорбируют ионы Na+ и Cl- из фильтрата (мочи) посредством Na+/К+/Cl--котранспортеров, находящихся в апикальной мембране,
и через CLC-каналы базолатеральной мембраны.