Ионы Са2+ представляют собой вторичный мессенджер, принимающий участие в многочисленных процессах передачи сигнала в различных клетках. У высших организмов внутриклеточный Са2+ участвует в таких разнообразных процессах, как синаптическая передача, мышечное сокращение, секреция инсулина, оплодотворение и экспрессия генов. В данной статье мы рассмотрим вопросы регуляции мышечного сокращения и сердечного ритма ионами Са2+.
Процесс, в результате которого деполяризация мембраны приводит к генерации мышечных усилий, называется сопряжение возбуждения и сокращения. Это фундаментальный механизм, контролирующий функции скелетных и сердечной мышц. В цитозоле клеток покоящихся мышц поддерживается гораздо более низкая концентрация свободного Са2+ (10-7 М), чем во внеклеточной среде (10-3 М) и в саркоплазматическом ретикулуме.
В начале процесса сопряжения возбуждения и сокращения ионы Са2+ начинают поступать в цитозоль, а по мере возвращения клетки в состояние покоя выходят оттуда. Такое увеличение и снижение концентрации Са2+ в цитозоле представляют собой фазный момент в процессе сопряжения возбуждения и сокращения. Для его протекания необходимо участие нескольких типов Са2+-транспортных белков.
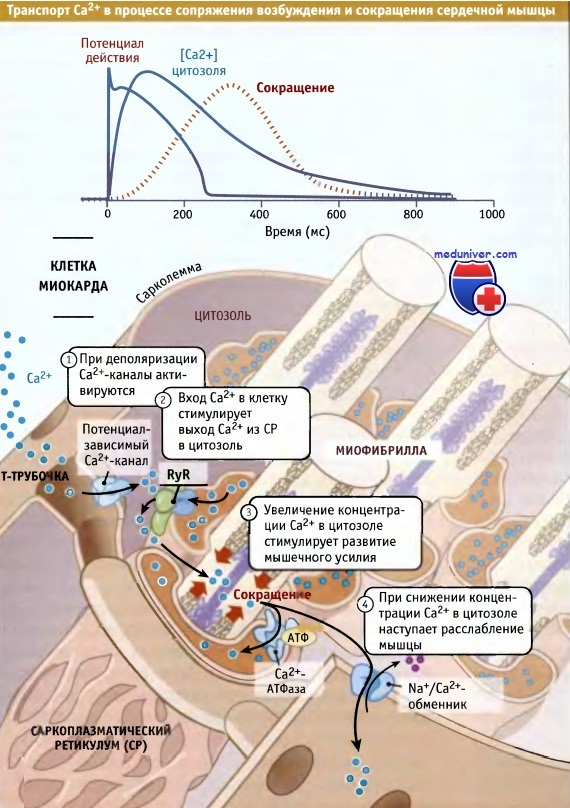
В клетках сердечной мышцы процесс возбуждения и сокращения подразделяется на четыре стадии.
На первой стадии, при деполяризации плазматической мембраны (сарколеммы) на ней генерируется электрический сигнал. При этом, благодаря входящему потенциалу действия, мембранный потенциал приобретает более положительное значение по сравнению с потенциалом покоя. Потенциал-зависимые Са2+-каналы (которые называются Cav1.2 Са2+-каналы) улавливают эти изменения мембранного потенциала и открываются (фаза 2 потенциала действия клеток миокарда). Небольшой поток ионов Са2+ начинает транспортироваться в клетку в направлении электрохимического градиента.
На второй стадии при поступлении Са2+ через Cav1.2 Са2+-каналы начинается высвобождение Са2+ из саркоплазматического ретикулума, в котором он депонирован в миллимолярных концентрациях. Выход Са2+ из саркоплазматического ретикулума происходит через особые каналы, называемые рианодиновые рецепторы (RyRs). В клетках сердечной мышцы этот процесс носит название Са2+-зависимый выход Са2+. Количество Са2+, выходящее в цитозоль из саркоплазматического ретикулума, в несколько раз превышает поступающее в цитозоль через сарколемму.
Са2+-зависимые Са2+-каналы саркоплазматического ретикулума клеток миокарда называются рианодиновыми рецепторами, поскольку они специфически связывают растительный алкалоид рианодин, блокирующий их действие. В различных клетках экспрессируются разные каналы внутриклеточного высвобождения Са2+, которые открываются в ответ на разнообразные сигналы, вызывающие мышечное сокращение. Основной вид каналов, высвобождающих Са2+ из саркоплазматического ретикулума клеток миокарда, относится к изоформе RyR2.
При генерации и распространении потенциала действия в клетках миокарда концентрация Са2+ в цитозоле увеличивается и снижается.
Это обеспечивается несколькими различными типами белков, транспортирующих Са2+.
На третьей стадии увеличение концентрации ионов Са2+ в цитозоле приводит к активации Са2+-зависимого белка тропонина С, который стимулирует сокращение мышечных волокон. Для эффективной активации всех внутриклеточных микрофиламентов и сокращения сердечной мышцы необходимо увеличение концентрации Са2+ в цитозоле со 100 нМ до 1,0 мкМ.
На четвертой стадии, когда Са2+ вытесняется из цитозоля, наступает расслабление мышцы. Вытеснение Са2+ происходит по нескольким механизмам. Основной из них — мобилизация Са2+ обратно в депо саркоплазматического ретикулума с помощью Са2+-АТФазы. Этот фермент осуществляет функцию насоса и направляет обратно в депо Са2+, который высвободился из саркоплазматического ретикулума при участии RyRs. Наряду с этим, Са2+ удаляется из цитозоля при участии таких Са2+- транспортных белков, как Na+/Са2+-обменник плазматической мембраны.
Этот обменник выводит из цитозоля небольшую часть Са2+, поступившего через потенциал-зависимые Cav1.2 Са2+-каналы. Немного Са2+ также обменивается между цитозолем и митохондриями.
В общем, за небольшими исключениями, процессы возбуждения и сокращения в скелетных и сердечной мышцах сходны. В отличие от клеток миокарда, потенциал-зависимые Са2+-каналы плазматической мембраны скелетных мышц представлены другой изоформой и стимулируют выход Са2+ из саркоплазматического ретикулума при физическом взаимодействии с изоформой рианодинового рецептора — RyRl. Наряду с этим, активация скелетных мышц может постепенно нарастать за счет вовлечения в процесс все большего количества мышечных волокон, что приводит к увеличению силы сокращения.
Таким образом, активация скелетных мышц может варьировать от коротких одиночных до повторяющихся тетанических сокращений и в конце концов ограничивается степенью мышечной усталости.
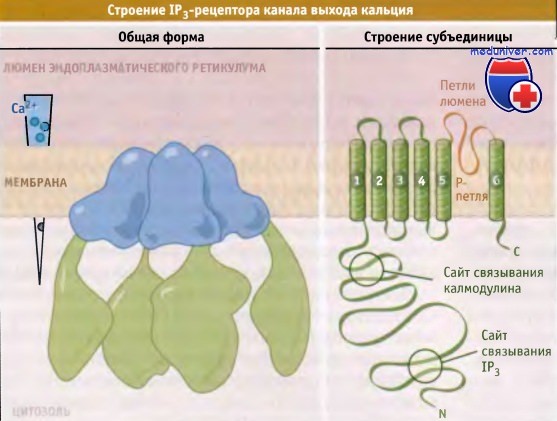
Предполагаемая структура инозитол-1,4,5-трифосфатного рецептора эндоплазматического ретикулума.
Слева показан Са2+-градиент, создающийся по сторонам мембраны эндоплазматического ретикулума в покоящихся клетках животных.
Каналы высвобождения внутриклеточного Са2+ относятся к числу уникальных ионных каналов. Они подразделяются на две группы: RyRs включаются и выключают ся под действием Са2+ или при прямом взаимодействии с Са2+-каналами плазматической мембраны, и близкие к ним рецепторы инозитол-1,4,5-трифосфата (IP3Rs), которые регулируются IР3. Четыре RyR или IP3R субъединицы собираются в симметрично организованный комплекс, образующий канал. Оба типа каналов состоят из двух доменов: порового и большого цитоплазматического, снабженного воротным механизмом. Предполагается, что канал IP3Rs имеет шесть трансмембранных сегментов и одну поровую петлю на субъединицу. Близкая структура постулируется и для RyRs.
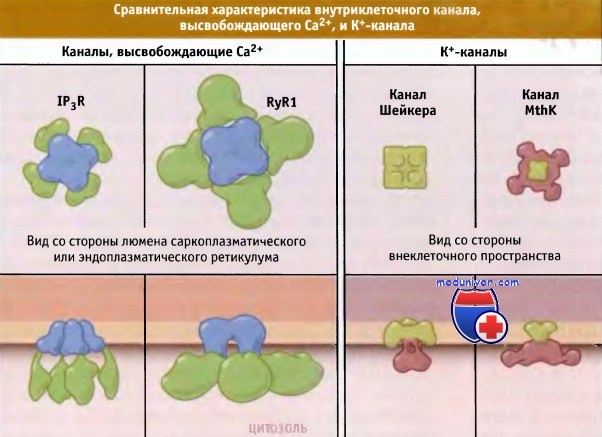
Са2+-каналы RyR представляют собой самые большие из известных ионных каналов. Структура их доменной организации получена при трехмерном моделировании с использованием данных электронно-микроскопических исследований. Эти каналы в 10 раз больше, чем Na+-, Са2+- или К+-каналы. Каждая субъединица RyR состоит примерно из 5000 остатков аминокислот, почти вдвое превышая размер частично гомологичной IP3R субъединицы. Величина поровых доменов RyRs и IP3Rs примерно такая же, как для К+-каналов. Большие цитоплазматические домены RyRs и IP3Rs контролируют открытие-закрытие каналов с помощью Са2+ и IР3, подобно тому, как это имеет место в лиганд-зависимых К+-каналах.
Мутации, нарушающие процессы открывания-закрывания каналов внутриклеточного высвобождения Са2+, приводят к развитию различных заболеваний. Например, миссенс-мутации в гене, кодирующем белок RyR2, связаны с двумя генетическими формами аритмии и внезапной смерти при физической нагрузке. Мутантная форма RyR Са2+-канала обладает пониженным сродством к калстабину 2 (который также называется FKBP12.6). Калстабин2 представляет собой субъединицу Са2+-канала, которая стабилизирует закрытое состояние RyR миокарда, предотвращая его аберрантную активацию.
В результате мутации в белке RyR в фазе покоя или диастолы сердца, из саркоплазматического ретикулума начинается усиленный выход ионов Са2+. Более того, разрегулирование RyR2 при болезнях сердца увеличивает вероятность ухудшения сердечной функции и наступления внезапной смерти. К общему механизму, вероятно, относится внутриклеточная утечка Са2+, которая за счет аберрантной деполяризации мембраны может вызвать угрожающую аритмию.
Мутации в гене, кодирующем RyR-изоформу клеток скелетных мышц, также приводят к аберрантному выходу внутриклеточного кальция, что служит причиной заболевания, известного под названием злокачественная гипертермия. Больные крайне чувствительны к неконтролируемому внутриклеточному выходу Са2+, при этом у них поднимается температура и отмечается генерализованная мышечная контрактура. При применении в ингаляциях некоторых анестетиков и мышечных релаксантов у больных могут наступить серьезные нарушения метаболических процессов, представляющие угрозу для жизни.
Мутации в гене, кодирующем белок потенциал-зависимых Са2+-каналов плазматических мембран скелетных мышц, который физически взаимодействует с RyRs и активирует его, также обеспечивают восприимчивость к развитию злокачественной гипертермии.
Два внутриклеточных канала, высвобождающих Са2+:
IР3-рецептор (IP3R) и рианодиновый рецептор (RyR), и два К+-канала: потенциал-зависимый канал Шейкера и Са2+-зависимый канал MthK различаются по размерам.
Поровые домены обозначены голубым и желтым цветом соответственно.