Механизм созревания эндосом и их превращения в лизосомы
• Транспорт метаболитов от ранних эндосом в поздние эндосомы и лизосомы происходит за счет «созревания»
• Белковые комплексы ESCRT участвуют в сортировке белков в везикулы, которые отпочковываются в люмен эндосом, образуя мультивезикулярные тела, обеспечивающие процесс протеолитической деградации
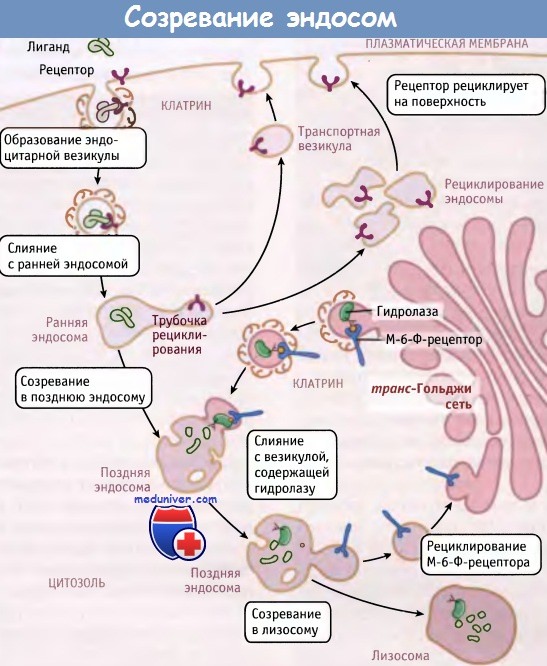
Превращение ранних эндосом в поздние эндосомы и в лизосомы происходит в ходе процесса созревания, который, по крайней мере, на первый взгляд напоминает созревание цистерн аппарата Гольджи. Вероятно, транспортные везикулы не играют существенной роли в движении белков между эндосомами.
Вместо этого потеря рецепторов за счет их возврата в плазматическую мембрану превращает ранние эндосомы в поздние эндосомы, содержащие диссоциированные лиганды, такие как ЛНП. Транспортные везикулы, содержащие вновь синтезированные ферменты лизосом и мембранные компоненты, происходят из транс-Гольджи сети и сливаются с поздними эндосомами. В ходе этого процесса поздние эндосомы превращаются в лизосомы, и происходит переваривание диссоциированных лигандов.
Образование цилиндрических отростков рециклизации ведет к селективному и постепенному удалению рецепторов из ранних эндосом. Остальные компоненты остаются в везикулярной части ранней эндосомы, которая перемещается в центр клетки вдоль микротрубочек. В конце концов везикулы накапливаются в перинуклеарной части цитоплазмы, примыкающей к центру организации микротрубочек. Одна из причин такого движения везикул заключается в необходимости ограничить их дальнейшее слияние с новыми клатриновыми везикулами, которые отпочковываются от плазматической мембраны.
Однако их слияние с везикулами, которые содержат лизосомальные компоненты и произошли из транс-Гольджи сети, может продолжаться. Таким образом, органеллы, принимающие участие в эндоцитозе, постепенно становятся более «лизосомоподобными» по составу содержащихся в них ферментов, Н+-ионов и мембранных компонентов, таких как v-АТФазы. Это превращение происходит постепенно, с образованием поздних эндосом как промежуточных структур между ранними эндосомами и лизосомами.
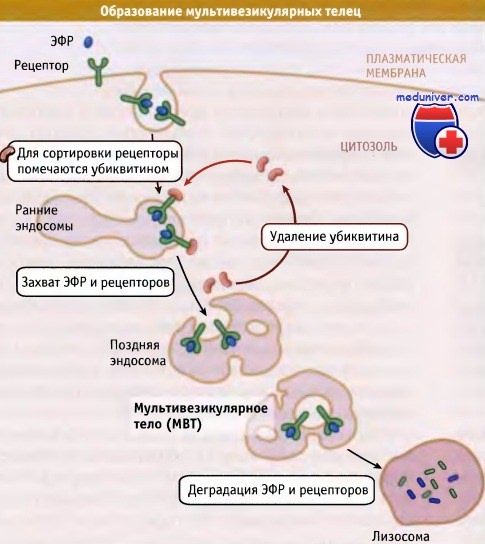
Характерная особенность процесса созревания ранних эндосом заключается в образовании мультивезикулярных тел (МВТ). В компартментах, принимающих участие на поздних этапах эндоцитоза (поздние эндосомы и лизосомы), часто присутствуют небольшие везикулярные включения, которые образуются в результате инвагинации мембраны. Таким образом, нередко поздние эндосомы и лизосомы называют МВТ. Функциональная роль этих структур особенно отчетливо проявляется в случае семейства рецепторов тирозинкиназ, например инсулинового рецептора и рецептора эпидермального фактора роста (ЭФР), которые после захвата лиганда деградируют.
Например, десенситизация ЭФР-рецептора происходит потому, что после попадания в ранние эндосомы рецептор селективно включается в участки мембран, которые находятся внутри эндосомы. Мембраны внутренних везикул по своим свойствам отличаются от мембран, ограничивающих эндосомы, и таким образом, рецепторы не попадают в цилиндрические отростки для последующего рециклирования. Вместо этого, совместно с диссоциированными лигандами и другим содержимым ранних эндосом, они поступают в поздние эндосомы и лизосомы, где внутренние везикулы вместе с накопленными рецепторами деградируют под действием липаз (ферментов, разрушающих липиды) и протеаз.
Исследования, проведенные на мутантах дрожжей, дефектных по образованию МВТ, позволили выяснить механизм их образования и отбора белков для мембран внутренних везикул. По-видимому, аналогичный механизм существует и для клеток млекопитающих. Характерной его чертой является метка подлежащих десенситизации рецепторов с помощью ковалентного присоединения к ним убиквитина, небольшого белка, присутствующего в цитоплазме. Молекула убиквитина присоединяется к рецептору с помощью специфических лигаз. Одна из таких лигаз представлена белком Cbl, который кодируется протоонкогеном и содержится в цитозоле. Этот белок катализирует присоединение одного убиквитина к рецептору ЭФР.
Дефект в белке Cbl приводит к нарушению процесса десенситизации ЭФР-рецептора и к неконтролируемому росту клеток. Эти моноубиквитинированные белки узнаются комплексом специальных белков, который обеспечивает их селективный захват во внутренние везикулы, а также участвует в образовании самих везикул. Поскольку эти события начинаются на уровне ранних эндосом, образование МВТ служит этапом на пути созревания ранних эндосом в поздние эндосомы и лизосомы. Исследования образования МВТ у дрожжей и в клетках животных показали, что один из белков цитозоля (Hrs) узнает остатки убиквитина и активирует три дополнительных набора белков, которые называются комплексами ESCRT. Эти комплексы удаляют убиквитин, обеспечивая его последующее использование, и, что более важно, способствуют образованию инвагинирующей внутрь мембранной везикулы с карго, с которого снят убиквинтин.
Важно отметить, что направленность формирования везикул при образовании МВТ имеет противоположный характер, по сравнению с направленностью всех известных на сегодня событий мембранного перемещения. Это объясняется тем, что целью образования МВТ служит выведение отобранных мембранных компонентов из транспортного пути, гарантирующее их последующее переваривание в лизосомах.
По мере созревания ранних эндосом в поздние,
транспортные везикулы доставляют к ним компоненты и рециклируют из них рецепторы.
Деградация белков с участием убиквитина происходит в мультивезикулярных тельцах,
представляющих собой поздние эндосомальные компартменты на пути эндоцитоза.