Охарактеризовано большинство белков, участвующих в процессах перемещения между мембранами, и в ряде случаев выяснены их функции. Однако о функциях многих белков можно только догадываться, и ощущается необходимость использования более углубленных методов исследований в этом направлении.
Например, в настоящее время удержание везикул на мембране рассматривается как событие в категории все или ничего, и упускается тот факт, что этот процесс очень тонко регулируется во времени и пространстве. По-видимому, наиболее адекватным в данном случае является сочетание биохимических и микроскопических методов исследования.
Более того, ощущается необходимость интеграции отдельных процессов для создания целостной картины. Часто можно изучить отдельные этапы, но путь, по которому они объединяются вместе, остается неясным. Примером является процесс выбора карго для транспорта в базолатеральную мембрану поляризованных клеток. Существует очевидная необходимость в специфическом адаптере (АР-1В), но как он связан с процессом сборки окаймления?
Адаптеры, которые связываются с базолатеральным карго и с М-6-Ф рецепторами, оба связываются с клатрином, но для них характерно различное направление. Что препятствует вхождению обоих типов комплексов адаптер-карго в одни и те же транспортные везикулы? Что регулирует и координирует индивидуальные события мембранного транспорта в процессе биологического развития или при метаболических изменениях в клетке или в тканях? Наблюдение и анализ таких событий в реальном времени возможны с помощью биохимических и микроскопических методов.
С другой стороны, существуют сложные структуры, которые исследуются методами рентгеноструктурного анализа и ядерного магнитного резонанса. Например, недавно получена структура адаптерного комплекса АР-2. Выяснение этих структур обеспечивает необходимые предпосылки для дальнейшего изучения механизмов процессов с использованием генетических и биохимических подходов.
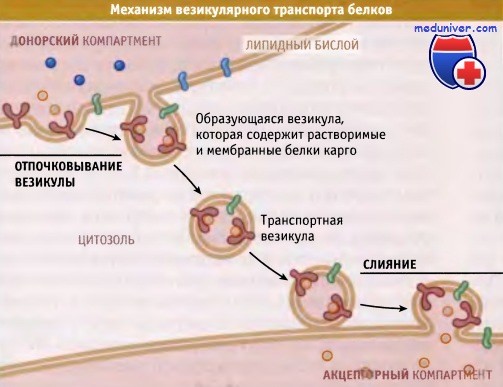
При везикулярном транспорте везикула отпочковывается
от мембраны одного компартмента и сливается с мембраной другого компартмента.
Уникальный набор белков отражает специализированные функции органелл, участвующих в экзо- и эндоцитозе. Состав этого набора поддерживается постоянным, несмотря на большое количество белков, которые проходят через органеллы в ходе их функционирования и в результате биосинтетических процессов. Индивидуальность состава компартментов поддерживается постоянным обменом везикулами между мембранами. Белки селективно включаются в везикулы, которые переносят их к следующему компартменту, и затем везикулы сливаются только с этим компартментом.
Транспортируемые белки сортируются в соответствии с сигналом, который прямо или опосредованно связывается с белком, окаймляющим везикулу. Этот механизм особенно характерен для эндоцитоза и в меньшей степени для эзоцитоза. Последний характеризуется продвижением молекул карго, особенно синтезируемых в больших количествах, или больших структур в виде неизбирательного потока по направлению к транс-Гольджи сети. Прямой селекции белков не происходит, и основную роль играют два других сигнала: удержание и возвращение.
За счет первого белок удерживается на специфической мембране по механизму, в котором участвует домен, пронизывающий мембрану. Второй сигнал обеспечивает захват белка и его возвращение в органеллу, где он должен функционировать. Многие растворимые и присутствующие в больших количествах белки ЭПР обладают С-терминальным сигналом KDEL, который служит для возвращения белков из ранних компартментов Гольджи. Для белков мембран ЭПР характерен сигнал, состоящий из остатков основных аминокислот.
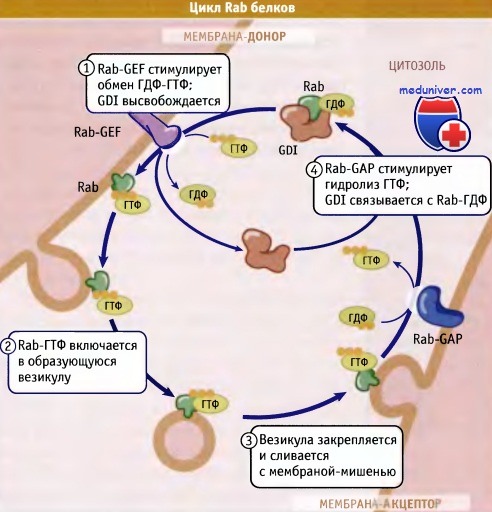
Для направления везикул к определенным компартментам необходимы Rab-ГТФазы, удерживающие белки и белки SNARE. Парные SNARE взаимодействуют между собой, при этом мембраны соприкасаются и сливаются. Слияние мембран контролируется, и таким образом регулируется секреция многих типов карго — от пищеварительных ферментов до нейромедиаторов.
Модель, описывающая цикл Rab-белков между цитозолем и мембранами
(другие белки, участвующие в перемещении, не показаны).