• Реснички и жгутики содержат высокоупорядоченную структуру, называемую аксонемой
• Аксонема состоит из девяти наружных дуплетов микротрубочек, окружающих пару центральных микротрубочек
• Радиальные спицы представляют собой комплекс из нескольких полипептидов, которые связывают каждый наружный дуплет с центром аксонемы
• С каждым внешним дуплетом связываются динеины, домены моторов которых распространяются на примыкающие внешние дуплеты
• Динеин сдвигает внешние дуплеты по отношению друг к другу; структурные связи между другими дуплетами переводят скольжение в наклон аксонемы
• Кинезины участвуют в сборке жгутиков, транспортируя белки аксонемы в отдаленный конец жгутика
• Неподвижные первичные реснички участвуют в сенсорных процессах
Наряду с транспортировкой карго в клетках, микротрубочки также участвуют в перемещении клеток в окружающей среде. Это перемещение осуществляется с помощью ресничек и жгутиков, длинных и тонких выростов, которые отходят как волоски на поверхности многих клеток. Каждая такая органелла состоит из длинного пучка микротрубочек, окруженного выростом плазматической мембраны. Взаимодействие между микротрубочками вызывает изгиб структуры, который возможен в разных направлениях. В результате этих движений жидкость может продвигаться вдоль поверхности клеток, как показано на рисунке ниже. Для группы неподвижных клеток, например эпителиальных, это позволяет жидкой среде и различным объектам перемещаться вдоль поверхности ткани. В случае изолированных, неприкрепленных клеток они сами движутся в жидкости (т. е. плавают). Реснички и жгутики присутствуют у многих одноклеточных организмов, таких как Paramecium и Chlamydomonas (зеленая водоросль), а также находятся на сперматозоидах у большинства эукариот. У млекопитающих реснички покрывают апикальные участки некоторых эпителиальных клеток и совершают синхронные движения. При этом возникают волны движения ресничек, которые проходят по поверхности ткани. В трахее это движение используется для удаления слизи и дебриса из дыхательных путей; в яйцеводе оно обеспечивает транспортировку яйцеклетки из яичника в матку, а в мозгу создает циркуляцию цереброспинальной жидкости.
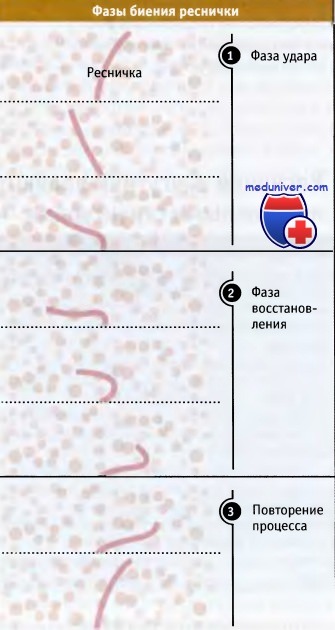
Реснички и жгутики обладают одинаковой структурой и механизмом подвижности, однако в некоторых отношениях они различаются. Наиболее существенные различия касаются длины, их количества в клетке и характера движения. Реснички короче (10-15 мкм) и количество их на клетку достигает 100 или больше. Каждая ресничка генерирует усилие за счет наклона у основания. Внешняя часть реснички остается жесткой и перегиб у основания сдвигает ее в движение, напоминающее гребок весла в воде. После этого наступает восстановительный размах, при котором перегиб реснички распространяется от основания на конец, подготавливая ресничку к следующему циклу движения. На рисунке ниже представлено движение реснички Для того чтобы проследить за отдельными стадиями движения, для регистрации использовали замедленную видеосъемку. Движения реснички столь интенсивны (много наклонов в секунду), что при съемке в обычном режиме она выглядит нечетко.
Жгутики обычно длиннее (10-200 мкм), чем реснички, и обычно клетка содержит только один или два жгутика. Они также генерируют силу при изгибе; при этом, как показано на рисунке ниже, от основания к вершине жгутика распространяется S-образная волна. Для ресничек и жгутиков характерен общий механизм движения, в основе которого лежит изгибание структуры. Различные пути распространения изгиба вдоль органеллы обусловливают различные формы волн, генерируемых ресничками и жгутиками. Поскольку два типа органелл представляют собой вариации на одну и ту же тему, мы рассмотрим их общие свойства и будем пользоваться термином жгутик для описания структуры и подвижности обеих органелл, если специально не оговаривается, что речь идет о ресничках.
При отделении от клетки жгутики продолжают биение. Это говорит о том, что оно обеспечивается самой органеллой. После удаления плазматической мембраны изолированные жгутики также продолжают биться при условии, что у них сохранился запас АТФ. Эти данные свидетельствуют о том, что движения обеспечиваются белковым содержимым жгутика и энергией гидролиза АТФ.
Внутренний белковый компонент жгутика представляет собой упорядоченную структуру, состоящую по крайней мере из 250 различных полипептидов. Эта структура называется аксонемой. Структура аксонемы жгутиков одинакова для столь различных организмов, как одноклеточное простейшее Chlamydomonas и человек.
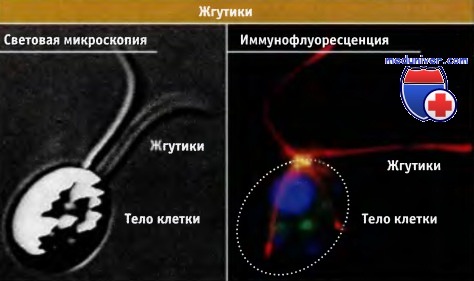
Одноклеточная водоросль Chlamydomonas reinhardtii в световом и флуоресцентном микроскопах.
В верхней части клетки видны два больших жгутика.
Во флуоресцентном микроскопе микротрубочки флуоресцируют красным; показывая, что жгутики состоят из микротрубочек.
Клетки Chlamydomonas способны передвигаться в воде за счет биений жгутиков.
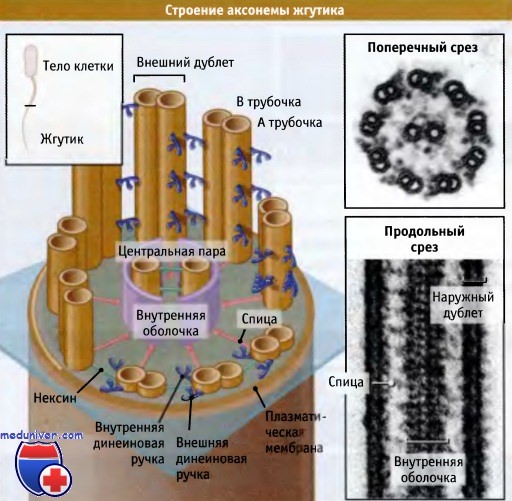
Строение аксонемы представлено на рисунке ниже. Наиболее примечательным элементом структуры, особенно заметным на поперечном срезе, являются определенным образом организованные дуплеты микротрубочки, расположенные вдоль всей аксонемы. Девять необычных «двойных микротрубочек» расположены по кругу. Каждая пара состоит из одной обычной микротрубочки, содержащей 13 протофиламентов (т. н. А трубочка) и одной неполной микротрубочки (В трубочка). Эта микротрубочка состоит из 10-11 протофиламентов и связана со стенкой А трубочки. В центре кольца, образованного дуплетами микротрубочек, находятся две обычных микротрубочки, содержащие 13 протофиламентов («центральная пара»). Такое расположение микротрубочек в аксонеме сокращенно описывается как «9+2». Все микротрубочки обладают одинаковой полярностью, их плюс-концы направлены к кончику жгутика, а минус-концы — к его основанию. С микротрубочками связываются различные белки, оказывающие на них стабилизирующее действие.
В аксонеме микротрубочки связаны между собой множеством различного типа связей. Белки, которые формируют эти связи, необходимы для организации микротрубочек в единую согласованную систему, способную двигаться и координировать эти движения для придания им волнового характера. Соседние дуплеты микротрубочек соединены между собой по периферии аксонемы белком, называемым нексином. Дуплеты микротрубочек также связаны с микротрубочками, расположенными в центре, с помощью полипептидных комплексов, которые образуют как бы спицы с видимыми головками. Эти структуры имеют сложное строение: спицы и их головки содержат 17 различных полипептидов. Головки расположены вокруг внутренней оболочки, структуры, окружающей две центральные микротрубочки. Усилие в аксонеме генерируется с помощью аксонемных динеинов (их также называют «ресничатыми» или «жгутиковыми» динеинами). Эти белки связывают между собой соседние дуплеты микротрубочек; хвостовой домен связывается с А трубочкой в одного дуплета, а головной домен с В трубочкой следующего дуплета. Различные связи, образованные нексином, спицами и динеинами, расположены через правильные промежутки по всей длине аксонемы, однако они характеризуются разной периодичностью. Поэтому все три компонента трудно наблюдать в электронном микроскопе на препаратах, представляющих собой поперечные срезы аксонем. Однако, когда удается видеть все три структурных элемента, они напоминают колесо с массивными спицами и втулкой.
Структура и расположение динеинов в остальной части аксонемы представляются сложными. В аксонеме содержится более одной формы динеина, каждая из них достаточно велика и состоит из большего числа различных полипептидов по сравнению с динеином, содержащимся в цитоплазме. Различные формы содержат один, два иди три моторных домена, и расположены в разных местах аксонемы. Смежные дуплеты микротрубочки связаны двумя наборами молекул динеина, которые называются внутренними и внешними ручками. Внешние ручки содержат только динеины с двумя или тремя головками, а белки с одной или двумя головками находятся во внутренних ручках.
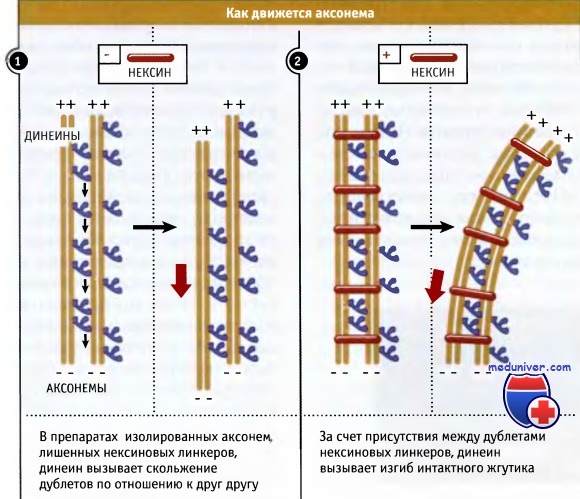
Каким образом все эти связи обеспечивают подвижность жгутиков и задают соответствующий характер биения? Основной вопрос заключается в том, как функционируют динеины в этой структуре, поскольку они представляют собой моторы и с них должно начинаться движение. Для того чтобы выяснить роль динеина в обеспечении подвижности, необходимо отделить жгутики от клеток и удалить мембрану аксонемы. После этого аксонемы подвергают кратковременной обработке протеазой для того, чтобы разрушить нексиновые связи между внешними дуплетами микротрубочек. Если затем к препарату аксонем добавить АТФ, то микротрубочки сдвигаются по отношению к друг другу. Этот эффект вызывается динеинами, хвосты которых связаны с дублетом микротрубочек, генерирующих усилие в направлении от плюс- к минус-концу соседней пары. В интактной аксонеме динеин не может сдвигать внешние дублеты, поскольку они связаны между собой с помощью нексина. Поэтому усилие, генерируемое динеином, приводит к сгибанию жгутика.
Реснички и жгутики генерируют биение за счет распространения сгибательного движения по аксонеме. Сгибание начинается в основании реснички или жгутика и распространяется по направлению к дистальному концу. Сгибание происходит потому, что динеин активен только в пределах небольшого участка аксо-немы. Для того чтобы обеспечить сгибание, динеины активируются последовательно как вдоль длины, так и по периферии аксонемы. Активность динеинов контролируется центральными микротрубочками и спицами; жгутики мутантных клеток, у которых эти структуры отсутствуют, не способны совершать биение. У некоторых организмов центральные микротрубочки быстро вращаются, и в это время они могут передавать сигналы на спицы, которые, в свою очередь, активируют динеин. В центральных микротрубочках и в спицах находится несколько киназ и фосфатаз. Предполагается, что при вращении центральных трубочек запускается локальная система передачи сигнала, которая активирует динеины. Посредством быстрой локальной активации и инактивации специфических изоформ динеина аксонемы генерируют движения и регулируют силу и частоту биений.
В основании жгутика находится структура, которая называется базальным тельцем. Эти структуры обладают таким же строением, как и центриоли. Каждое базальное тело представляет собой цилиндр, состоящий из 9 триплетов микротрубочек, каждая из которых состоит или из 13 (трубочка А) или из 11 протофиламентов (трубочки В и С). Трубочки А и В служат матрицами при сборке 9 наружных дуплетов микротрубочек аксонемы. Базальное тело остается связанным с основанием аксонемы и служит для прикрепления последней к телу клетки.
Как происходит сборка жгутика, можно выяснить, удалив его с поверхности клетки и проследив образование нового. Жгутик регенерирует менее чем за час и в ходе этого процесса функционирует (т. е. осуществляет биения). Рост нового жгутика происходит на плюс-концах микротрубочек аксонемы, расположенных в дистальных участках каждого жгутика. Сборка жгутика предполагает, что необходимые компоненты аксонемы должны транспортироваться на концы структур и по мере роста собираться в аксонемы. Транспорт осуществляется в больших белковых комплексах, которые движутся в направлении кончика жгутика по наружной поверхности аксонемы, расположенной непосредственно под плазматической мембраной. Это движение называется интрафлагеллярным транспортом (IFT) и осуществляется с участием кинезина. Белковые комплексы также могут двигаться от вершины жгутика к его основанию (к минус-концам микротрубочек), однако функциональное значение этого направления транспорта неизвестно. IFT по направлению к телу клетки происходит за счет цитоплазматического динеина.
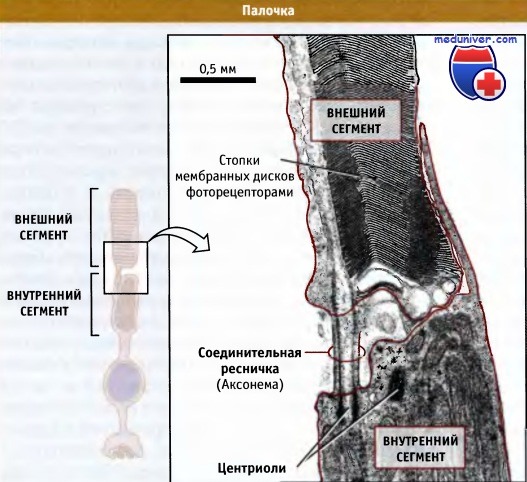
Хотя большинство ресничек представляют собой подвижные образования, существуют формы этих структур, которые не обладают подвижностью и играют в клетках особую роль. Первичные реснички представляют собой неподвижные органеллы, обнаруженные практически во всех клетках позвоночных, исключая клетки крови. В отличие от подвижных ресничек, у клеток обычно находится только одна неподвижная ресничка. На рисунке ниже представлен наиболее типичный пример. Аксонема первичной реснички не содержит центральных микротрубочек, и поэтому ее структура часто обозначается как «9+0». Снаружи большинство первичных ресничек выглядят как обычные реснички. Они начинаются на поверхности клеток и имеют вид простого короткого волоска. Однако, у некоторых высокодифференцированных типов клеток дистальный участок первичной реснички сильно вытянут и превратился в специальный домен, размеры которого сравнимы с размерами самой клетки. Это, например, имеет место в клетках палочек и колбочек, которые содержат фоторецепторы, участвующие в восприятии света сетчаткой глаза. В палочках кончики ресничек вытянуты и образуют большой домен, называемый наружным сегментом, который содержит стопки мембранных дисков, заполненные белковым фоторецептором, родопсином. На рисунке ниже представлен соответствующий пример. Основание первичной реснички соединяет наружный сегмент с остальной частью клетки, аксонема лишь немного вдается в область, где начинается наружный сегмент. За счет транспорта IFT типа, мембранные везикулы, содержащие родопсин, перемещаются из тела клетки в наружный сегмент, и вероятно, это необходимо для его образования и функционирования.
Использование наружного сегмента палочки в качестве светового сенсора может иллюстрировать широкое распространение такой возможности среди первичных ресничек. Возможно, что подобная функция первичных ресничек только начинает исследоваться. В других типах клеток содержатся более примитивные первичные реснички, в которых могут быть локализованы различные типы рецепторов. Локализация рецепторов в первичных ресничках может превратить их в некое подобие антенн, которые могут улавливать изменения в окружении и передавать эту информацию в клетку.
Некоторые редкие болезни человека связаны с мутациями, в результате которых реснички и жгутики утрачивают подвижность. Больные, которым эти мутации передаются по наследству, обычно страдают хроническими респираторными инфекциями, поскольку неподвижные жгутики не способны выводить из дыхательных путей слизь, а также захваченные патогенные микроорганизмы и раздражители. Больные мужчины часто оказываются бесплодными, так как сперматозоиды у них лишены подвижности. К числу широко известных заболеваний, связанных с неподвижностью ресничек и жгутиков, относится синдром Картагенера. Наряду с респираторными инфекциями и бесплодием, у половины больных отмечается извращенное расположение внутренних органов, при котором изменяется нормальная левосторонняя их асимметрия. Предполагается, что на одном из этапов раннего развития, до момента закладки внутренних органов, в результате биения жгутиков, в эмбрионе создается циркуляция жидкой среды, и возникает градиент секретирующих-ся веществ, способствующих морфогенезу, что определяет левостороннюю асимметрию. В отсутствие морфогенного градиента органы занимают случайное положение слева и справа. У мышей с мутациями по флагеллярным динеинам или моторам ответственным за IFT, также отмечается извращенное расположение внутренних органов. Это говорит о том, что мутации, влияющие на подвижность жгутиков или на их образование, могут приводить к порокам развития.
Биение реснички подразделяется на две фазы.
В момент удара ресничка полностью выпрямляется и проталкивает жидкость вдоль поверхности клетки.
В следующей затем фазе восстановления ресничка сгибается и возвращается в исходное положение для следующего удара.
Процесс биения реснички, наблюдаемый под микроскопом.
Отмечается резкий перегиб реснички у основания, происходящий в фазе удара и ее постепенное разгибание при восстановлении.
Процесс биения жгутика под микроскопом.
Строение аксонемы. Показано упорядоченное расположение микротрубочек.
Микротрубочки соединены между собой различными связями, согласованное действие которых обеспечивает биение реснички.
Справа представлены электронные микрофотографии. На поперечном срезе видны внутренние и внешние динеиновые ручки,
связывающие внешние дублеты микротрубочек. Также видна одна из спиц с головкой (внизу слева).
На кадрах анимации представлены два внешних дуплета (выделены золотистым цветом), соединенные динеином.
Первая часть анимации показывает, что происходит, когда из жгутиков выделяют дуплеты и селективно удаляют нексиновые линкеры.
Вторая часть показывает, что происходит в интактном жгутике. Под действием динеина и в присутствии нексиновых линкеров между дуплетами происходит перегиб жгутика.
Очень длинная первичная ресничка, отрастающая от клеточной поверхности.
Внизу представлены клетки в поперечном сечении; мембраны показаны пунктиром.
Вздутие на ресничке может представлять собой карго, которое транспортируется между аксонемой и окружающей мембраной.
Слева изображена зрительная палочка.
Показаны внутренний и внешний сегменты и тонкая перемычка между ними.
Справа представлена микрофотография области между двумя сегментами (выделенная на рисунке), сделанная в электронном микроскопе.
На небольшом расстоянии, вблизи от места, где она выходит из внутреннего сегмента,
перемычка выглядит как нормальная ресничка. Однако ее кончик погружен во внешний сегмент.