• Часто в процессе прикрепления хромосомы к веретену происходят случайные ошибки, которые исправляются
• Ошибочные прикрепления неустойчивы, поскольку они не позволяют кинетохорам стабилизировать прикрепившиеся микротрубочки
• Только правильное биполярное прикрепление хромосомы приводит к стабильному прикреплению кинетохора
Чувствительность места контакта микротрубочки и кинетохора к натяжению играет важную роль в коррекции ошибок прикрепления, которые происходят при формировании веретена. Механизм поиска-захвата может служить источником двух типов ошибок прикрепления, которые могут происходить в ходе процесса нормального митоза.
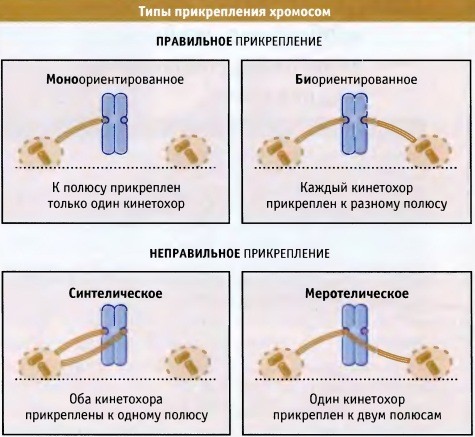
Хромосома прикрепляется синтелически (от греч. syn — тот же самый; telos — конец), когда оба ее кинетохора прикрепляются к одному и тому же полюсу. Ошибка обычно происходит сразу после разрушения оболочки ядра, когда хромосомы находятся в случайной ориентации и могут оказаться гораздо ближе к одной центросоме, чем к другой. Это дает возможность сестринским кинетохорам одновременно захватывать микротрубочки, исходящие из одной и той же центросомы.
Один кинетохор может прикрепляться сразу к двум полюсам (меротелическое прикрепление; греч. mero — часть). Хромосомы, содержащие один (или даже два) меротелически ориентированных кинетохора, нормально движутся к экватору веретена и могут представлять серьезную опасность для клетки. Если меротелическое прикрепление сохраняется, две хроматиды разделяются при вступлении клетки в анафазу, но ошибочно прикрепившаяся хроматида остается у экватора веретена.
Она находится там до тех пор, пока не нарушится ее связь с одним из полюсов, что обеспечит 50%-ю вероятность ее сегрегации к тому же полюсу, что и сестринская хроматида.
Биориентация является единственным типом прикрепления, который обеспечивает одинаковое распределение хромосом по двум дочерним клеткам;
все хромосомы должны быть биориентированы до начала анафазы.
Моноориентация представлет собой нормальный промежуточный этап на пути к биориентации.
В ранней прометафазе часто наблюдается синтелическое и меротелическое прикрепление,
однако до наступления метафазы эти типы превращаются в биориентированное прикрепление.
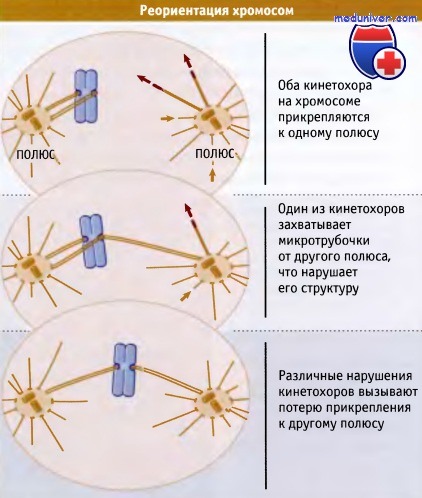
Обычно синтелические и меротелические прикрепления исправляются вскоре после того, как они образовались. Для обоих типов ошибочных прикреплений характерно, что нить кинетохора расположена не под прямым, а под острым углом к кинетохору. Это приводит к повреждению структуры кинетохора и к дестабилизации его связи с пучком микротрубочек.
В результате микротрубочки начинают отделяться быстрее, а замещаться медленнее, чем при правильном прикреплении. Это приводит к тому, что в конце концов в нити кинетохора больше не остается микротрубочек, и связь разрушается. В зависимости от типа ошибочного прикрепления, утрата связи приводит или к исправлению ошибки за счет создания биориентированной хромосомы, или к появлению хромосомы, прикрепленной только к одному полюсу через один кинетохор. Такая моноориентированной хромосома может затем по обычному механизму превратиться в биориентированную.
Важную роль в исправлении ошибочных прикреплений также играет чувствительность кинетохоров к натяжению. Ошибки прикрепления не позволяют кинетохорам обращаться к полюсу, тем самым предотвращая развитие натяжения, необходимого для их правильного прикрепления. Особенно характерна нестабильность для синтелического прикрепления, поскольку в этом случае на два кинетохора приходится незначительное натяжение, существенно меньшее, чем при правильной ориентации хромосомы и локализации центромеры между двумя полюсами.
Такая ошибка часто исправляется или при спонтанной потере одной из связей из-за отсутствия натяжения, или когда один из двух кинетохоров дополнительно прикрепляется к отдаленному полюсу. При этом в направлении полюса развивается резкое усилие, которое нарушает структуру кинетохора и дестабилизирует его прикрепление.
Чрезвычайная запутанность механизма прикрепления демонстрирует всю структурно-функциональную сложность кинетохоров и их центральное место в процессе митоза. Фактически расхождение реплицированных хромосом на две одинаковые группы непосредственно связано с функцией кинетохоров. В 1961 г. Мэзия — один из пионеров исследования механизмов митоза — обратил внимание на их исключительную роль. Он писал, что кинетохор «представляет собой единственную существенную часть митотической хромосомы».
Мэзия сравнивал остальную часть хромосомы с «трупом на похоронах» поскольку она «служит причиной события, но не принимает в нем непосредственного участия». Он был прав, считая, что истинная цель митоза состоит в сегрегации сестринских кинетохоров (остальную часть хромосомы можно рассматривать лишь как ездовые аксессуары). Однако в то время он не мог предвидеть, насколько глубоко вовлечены в этот процесс кинетохоры.
Итак, нити кинетохора прикрепляют хромосому к веретену и определяют направление ее движения. Одинаковая сегрегация хромосом в митозе может быть непосредственно связана со следующими установленными фактами:
(1) каждая реплицированная хромосома содержит два сестринских кинетохора;
(2) эти кинетохоры расположены на противоположных сторонах первичной перетяжки и обращены в противоположных направлениях;
(3) кинетохорные микротрубочки связываются только с его внешней частью, направленной в сторону от хромосомы: и
(4) существуют механизмы исправления ошибок прикрепления кинетохоров к полюсам. Отсутствие этих механизмов приводит к анеуплоидии.
Возможность исправления неправильного прикрепления хромосомы
зависит от динамических свойств микротрубочек в митозе и от чувствительности кинетохоров к возникновению у них различных нарушений.
Только при достижении конфигурации, изображенной внизу, может сформироваться нормальная нить кинетохора, и хромосома надежно связывается с веретеном.