• Хромосомы выстраиваются в метафазе под действием баланса нескольких сил
• В этом процессе участвуют силы, локализованные на кинетохорах и вдоль плечей хромосом
• Согласно наиболее вероятной модели, силы, направленные к полюсам, пропорциональны длине каждой нити кинетохора, позиционирующей хромосомы в центре веретена
• Этот механизм может выстраивать хромосомы в некоторых клетках
• Во многих типах клеток должны участвовать другие силы, включая те, которые генерируются кинетохором, а также отталкивающие хромосомы от полюсов
В 1945 г. Остергрен — еще один пионер в области исследований механизмов митоза — предложил изящное объяснение феномену конгрессии. Он предположил, что «положение равновесия (хромосомы) убедительно говорит о том, что центромеры (кинетохоры) притягиваются к полюсам веретена силами, увеличивающимися с увеличением расстояния между центромерой и полюсом и что каждая центромера притягивается только к тому полюсу, к которому она повернута».
Иными словами, величина силы, продвигающей хромосому к полюсу, которая действует на каждый кинетохор, должна быть пропорциональна длине ассоциированной с ним нити. По мнению Остергрена, хромосома движется к экватору веретена, потому что он является тем местом, где противоположно направленные нити кинетохора имеют одинаковую длину, и, таким образом, баланс сил, действующих на хромосому, равен нулю.
В пользу предложенной Остергреном модели «тянущей нити» свидетельствует наличие движения (потока) субъединиц тубулина по нити кинетохора. Поток предполагает наличие молекулярных моторов, закрепленных в матриксе веретена, который окружает микротрубочки. Эти моторы должны протягивать тубулиновые субъединицы по всей длине микротрубочек в направлении полюса. Результирующее движение микротрубочек к полюсу и представляет собой то, что мы называем поток.
Если такой механизм действительно существует, то, как предположил Остергрен, чем длиннее микротрубочка, тем больше будет сила продвижения, действующая на кинетохор и направленная к полюсу. Согласно этой модели, кинетохор просто удерживается на конце движущейся нити и подтягивается к полюсу веретена.
В таких клетках, как сперматоциты насекомых, а также у некоторых растений поток субъединиц микротрубочек служит единственным механизмом продвижения хромосом к полюсам. Для этих клеток модель тянущей нити остается наиболее вероятным объяснением механизма конгрессии хромосом. Для остальных клеток, однако, некоторые выводы, вытекающие из этой модели, оказываются неприменимыми, и возможно, что она лишь частично отражает происходящие в них процессы. Например, в клетках позвоночных вклад потока при движении хромосомы к полюсу составляет только 30% от ее реальной скорости.
Остальные 70% обусловлены активностью со стороны кинетохора. На первый взгляд, эти данные не подтверждают модель, предложенную Остергреном для объяснения конфессии в клетках позвоночных. Однако величина скорости потока не содержит никакой информации о величине развивающегося усилия. Поэтому возможно, что в клетках позвоночных усилие, направленное к полюсу и генерируемое потоком, существенно превышает развиваемое моторами на кинетохоре.
Для клеток позвоночных модель необходимо модифицировать, поскольку очевидно, что в этих клетках силы, тянущие кинетохор, не являются единственными действующими на хромосомы. К такому выводу приводят наблюдения за хромосомами, прикрепленными только к одному полюсу Если бы положение хромосомы определялось исключительно тянущим усилием, приложенным к кинетохорам, как постулирует модель Остергрена, то следовало бы ожидать, что такие моноориентированные хромосомы все время продвигались бы к полюсу.
Вместо этого они часто останавливаются на расстоянии нескольких мкм от полюса. Это позволяет предполагать, что существует другая сила, которая отодвигает хромосомы от полюсов.
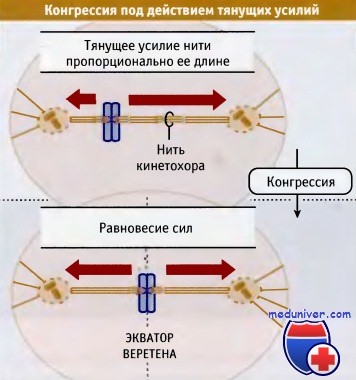
Механизм конгрессии по Остергрену.
Биориентированная хромосома движется слева направо, поскольку тянущее усилие,
приходящееся на каждый сестринский кинетохор, пропорционально длине его нити, причем более длинная нить обеспечивает большее усилие.
На рисунке длина и направление красных стрелок соответствуют величинам и направлению усилий.
Движение хромосомы продолжается до тех пор, пока две силы не уравновесятся.
Это наступает, когда хромосома оказывается на середине веретена.
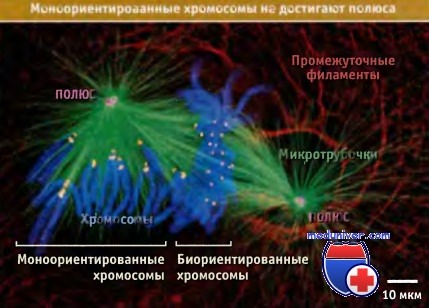
В этой прометафазной клетке примерно половина хромосом уже биориентирована и выстроена на середине веретена.
Остальные хромосомы (по крайней мере 7) еще находятся в моноориентации.
Хотя все моноориентированные хромосомы связаны только с левым полюсом, они располагаются от него на некотором расстоянии.
Желтые точки представляют собой кинетохоры. Отметим, что пары точек на биориентированных хромосомах все расположены параллельно оси веретена,
а точки на моноориентированных хромосомах по отношению к полюсу располагаются радиально.