После завершения проектов секвенирования генома были предприняты попытки приблизиться к пониманию характера изменений в генах и в функционировании их продуктов на протяжении клеточного цикла. Метод микроаррей-анализа позволил исследователям идентифицировать все изменения транскрипции, происходящие в клеточном геноме на протяжении цикла или после обработки клеток веществами, которые блокируют их продвижение по циклу.
Наиболее исчерпывающий анализ изменений профиля экспрессии генов выполнен на дрожжах S. cerevisiae и S. pombe. При этом были идентифицированы гены, транскрипция которых на протяжении клеточного цикла меняется. Аналогичным образом использовали микроаррей-анализ для идентификации генов, транскрипция которых при активации точки контроля увеличивается или снижается. Эту же стратегию также применяли к другим организмам для поиска общих принципов регуляции клеточного цикла и активации точек контроля, а также для оценки различий между здоровыми и трансформированными клетками животных.
Стратегия микроаррей-анализа сейчас используется для получения геномного профиля злокачественных клеток и выработки терапевтических подходов к лечению больных.
Еще один метод, который используется для идентификации регуляторных белков клеточного цикла, представляет собой глобальный анализ локализации белка. Например, была установлена локализация почти каждого белка, закодированного в геноме клеток S. cerevisiae и S. pombe. При этом использовался метод слияния каждой открытой рамки считывания с последовательностью, кодирующей зеленый флуоресцентный белок Также в перспективе намечается проведение крупномасштабных анализов с использованием методов протеомики, которые позволят определить белковые компоненты всех мультимерных комплексов, обеспечивающих жизнедеятельность клетки. Такой анализ мог бы способствовать нашему пониманию функционирования различных белков и их комплексов.
Принимая во внимание всю важность точек контроля в координации событий клеточного цикла, особенно необходимым представляется понимание молекулярных механизмов их функционирования. Точки контроля за повреждениями в ДНК, за ее репликацией в S-фазе, а также контролирующие сборку веретена, требуют участия ряда протеинкиназ. Однако пока идентифицировано лишь ограниченное число субстратов этих киназ, и представляется весьма важным продолжать поиск с тем, чтобы возможно более полно выяснить роль различных реакций фосфорилирования в процессах контроля. Выяснение всех перечисленных вопросов явилось бы крупным шагом вперед в понимании роли точек контроля в обеспечении точности передачи хромосом дочерним клеткам.
Аналогичным образом, несмотря на большое количество доступной информации относительно регуляции киназ митоза, предстоит еще многое выяснить о мишенях этих киназ. Совместное использование генетических методов на дрожжевых объектах и методов биохимического анализа при работе с экстрактами яйцеклеток лягушки и морского ежа обеспечило получение важных данных о функциях Cdk1 киназы митоза. Однако картина еще далека от завершения.
Как Cdk1 регулирует несколько таких митотических процессов, как конденсация хромосом, сборка веретена и разделение сестринских хроматид? Дальнейшая идентификация субстратов Cdk1 помогла бы полнее понять их молекулярные основы. Не менее важным представлется выяснить, каким образом выход клетки из митоза связан со снижением активности Cdk1. Как снижение активности фермента сопряжено с распадом веретена и цитокинезом? Вызваны ли эти процессы дефосфорилированием белков? Если да, то каким образом они регулируются путем дефосфорилирования белков?
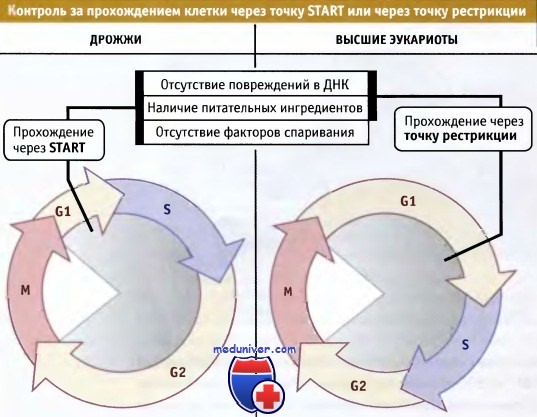
Прохождение клетки через точку START (у дрожжей) или точку рестрикции (у высших эукариот) служит сигналом о готовности клетки войти в цикл и регулируется внешними факторами,
такими как наличие необходимых для роста питательных ингредиентов или отсутствие гормонов спаривания (у дрожжей).
Процесс также регулируется внутренними факторами, например степенью интактности ДНК.
Постоянное усовершенствование масс-спектрометрических методов уже обеспечило простоту исследования таких модификаций белков, как фосфорилирование. Методы биохимической генетики оказались мощным инструментом в идентификации субстратов протеинкиназ. В этих методах используются ингибиторы модифицированных киназ для установления роли последних в специфических реакциях фосфорилирования. Такие аналог-чувствитель-ные аллели (аллели, содержащие мутантную форму киназы, которая не зависит от АТФ, но является аналогом дикой формы) оказались ценным инструментом для избирательного ингибирования этой специфической киназы и идентификации ее субстратов in vivo. Очевидно, что подобные методические подходы помогут исследователям ответить на следующие вопросы: Какие белки фосфорилируются на определенных стадиях клеточного цикла?
Насколько меняется степень фосфорилирования определенного белка после снижения активность определенной киназы? Ответы на все эти вопросы помогут составить полную картину того, как протеинкиназы регулируют критические процессы клеточного цикла.
В то время как протеинкиназы служат регуляторами критических событий клеточного цикла, направленный протеолиз обеспечивает необратимое продвижение клетки по циклу. SCF и АРС представляют собой две убиквитин-лигазы, специфические для клеточного цикла и необходимые для нормального протекания его процессов. Дефекты убиквитин-зависимого протеолиза также связаны с развитием опухолей. Поэтому идентификация белков, которые подвергаются убиквитин-зависимому протеолизу на протяжении цикла деления, играет критическую роль для понимания процессов клеточного цикла и развития опухолей.
Уже предприняты попытки обнаружить такие белки. Например, используя систему in vitro, в экстрактах яйцеклеток лягушки был идентифицирован набор белков, которые подвергаются специфическому протеолизу в митозе. Продолжение поиска таких белков и тщательный поиск субстатов SCF и АРС должны способствовать выяснению роли убиквитин-зависимого протеолиза в процессах цикла. Более того, выяснение деталей протеолиза может способствовать выявлению дополнительных мишеней для действия лекарственных средств.
Основную сложность в разработке средств химиотерапии рака представлял собой поиск агентов, селективно действующих на раковые клетки. Исследования всех тонкостей регуляции клеточного цикла должны дать в руки исследователям инструмент воздействия на регуляторные процессы, способные предотвратить наступление нежелательных циклов деления. Идентификация белков, участвующих в различных аспектах прохождения клетки по циклу, позволит составить список возможных мишеней для разработки противораковых средств. Аналогичным образом, поиск генов, участвующих в регуляции цикла, и использование метода генетических «отпечатков пальцев» помогут идентификации предрасположенности к определенному типу опухолей и оценке возможностей для фармакологического вмешательства.
Клеточный цикл представлет собой упорядоченную цепь событий, которые приводят к дупликации клеточного содержимого и к делению клетки на две. События клеточного цикла регулируются во времени и в пространстве. Определенный порядок и точность протекания процессов клеточного цикла обеспечиваются специальными механизмами мониторинга, известными под названием точек контроля. В случае возникновения дефектов процесса дупликации точки контроля задерживают прохождение клетки по циклу, обеспечивая возможность исправления ошибок.
Репликация генетической информации происходит в S-фазе, а сегрегация информации — в фазе митоза. Обе фазы разделены фазами-промежутками, G1 и G2. Такой порядок событий устанавливается точками контроля и реализуется с помощью основных регуляторных киназ клеточного цикла, CDKs. Некоторые процессы цикла наступают, только когда понижается активность Cdk1 (например, сборка пререпликативного комплекса), а другие — только при повышении ее активности (вступление в митоз). Походящие для репликации внешние условия устанавливаются, только когда активность Cdkl низкая, а митотический процесс начинается, когда активность киназы высока. Вхождение в фазу митоза завершается после полной активации Cdk1, а также некоторых других протеинкиназ. Эти киназы регулируют функцию машины сегрегации хромосом, а именно веретена.
После биполярного прикрепления сестринских хроматид к митотическому веретену при наступлении анафазы они расходятся и в дальнейшем мигрируют к противоположным полюсам веретена за счет его элонгации. Разделение сестринских хроматид в анафазе требует участия убиквитин-лигазы, АРС. Активность Cdk1 в анафазе начинает снижаться, что позволяет клетке входить в S-фазу следующего цикла.
Внешние сигналы определяют, должна ли здоровая клетка вступить в цикл деления или перейти в неделящееся, покоящееся состояние. Такие сигналы зависят от наличия питательных веществ, межклеточных взаимодействий, а также от присутствия ростовых факторов. Биохимические процессы, которые генерируются в ответ на поступление внешних сигналов, могут обладать стимулирующими или ингибиторными свойствами. В большинстве случаев, они влияют на точку перехода из G1- в S-период, которая для большинства клеток представляет собой основную точку контроля в цикле. Коммитирование клетки к вступлению в S-фазу, переход от G2 к митозу и наступление анафазы находятся под контролем соответствующих точек, которые задерживают прохождение клетки по циклу в случае, если не выполнены все требования, обеспечивающие точность прохождения событий цикла.
Дефекты механизма функционирования точек контроля приводят к сохранению ошибок дупликации в следующем поколении клеток и могут привести к нерегулируемой пролиферации и к развитию рака. Образованию опухолей способствуют два типа мутаций: инактивирующие мутации в генах-супрессорах опухоли и активирующие мутации в протоонкогенах. Одна мутация редко служит причиной возникновения опухоли, и риск развития рака возрастает, когда в клетке присутствуют несколько мутаций, нарушающих ее генетическую стабильность.