Расшифровка генома Arabidopsis thaliana служит стимулом к проведению дальнейших исследований в области биологии растительной клетки и процессов ее развития. Сейчас у растений можно идентифицировать гены, гомологичные хорошо известным генам дрожжей и клеток животных. Исследователи, изучающие клетки растений, вначале будут продолжать заниматься функциями белков в растениях. Однако не все белки растений имеют высокую степень гомологии с белками других эукариот, в том числе и с теми, которые могут выполнять различные функции.
Одна из трудностей состоит в том, что необходимо выяснить не только, где расположен тот или иной белок, но и как он функционирует, являясь компонентом комплекса. Важным примером такого рода служат исследования контроля направленности биосинтеза целлюлозы. Они не только включают характеристику всех ферментов, входящих в белковые комплексы синтеза целлюлозы и их ассоциации с мембранными липидами, но также выясняют то, как цитоскелет управляет движением комплексов. Существуют ли между ними прямые связи? С помощью современных методов возможно будет одновременно пометить синтезирующие комплексы и актин или микротрубочки. К тому же существует старый нерешенный вопрос: под влиянием каких факторов ориентируется цитоскелет? Используя методы анализа изображений, было показано, что синтазы целлюлозы движутся вдоль микротрубочек, но для того, чтобы решить, существует ли физическая связь между ними, необходимы дальнейшие исследования.
Еще один большой и сложный вопрос постгеномной эры состоит в том, чтобы понять, как собирается и функционирует клеточная стенка. При образовании большинства полисахаридных комплексов клеточной стенки моносахариды, могут полимеризоваться и образовывать разветвленные цепи фактически при неопределенно большом количестве комбинаций. Мы надеемся выяснить, не только как вначале синтезируются полисахариды и потом взаимодействуют между собой образуя внеклеточный матрикс, но и каким образом разные клеточные стенки — и даже различные стороны стенки в одной клетке — получают разные наборы полисахаридов.
Исследование процессов, контролирующих образование плоскости клеточного деления, относится к смежной области, находящейся между биологией клетки и биологией развития. Каким образом препрофазное кольцо определяет плоскость деления? Оставляет ли оно после себя некую «молекулярную зарубку» (такую как высокая концентрация белков, связывающихся с цитоскелетом, ассоциированным с плазматической мембраной), которая узнается цитоскелетом, связанным с фрагмопластом, растущим к периферии клетки? Однако вне вопроса о том, как определяется расположение плоскости деления в одной клетке, существует более сложный вопрос, касающийся координации плоскостей деления в группе клеток. Определяется ли расположение плоскости деления такими факторами морфогенеза, как гормоны, которые могут действовать вне границ отдельных клеток? Могут ли участвовать в этом физические силы, передающиеся в тканях на большие расстояния?
Одним из больших пробелов в наших знаниях является недостаток представлений о механизме действия гормонов растений. Необходимо выяснить, как регулируется баланс между делением и увеличением размера клеток меристемы и каким образом устанавливается гормональный контроль над повторным вступлением зрелых клеток в деление (например, после получения растением повреждений). Мы также не знаем, как воздействия со стороны таких внешних факторов, как свет и гравитация, транслируются в изменения роста и формы растений. По-видимому, изменение цитоскелета происходит где-то между приемом внешнего сигнала и морфологическим ответом, и, вероятно, мы лучше поймем это, когда больше узнаем о цепях передачи сигнала, связывающих первичный эффект гормонов с изменением цитоскелета. По-видимому, в этих процессах участвуют разнообразные пути передачи сигнала.
Клетки организмов растений и животных растут и функционируют совершенно по-разному. Различия между ними являются следствием наличия прочной и жесткой стенки, окружающей каждую клетку растения и не дающей им возможности двигаться или менять свою форму. Клетки растений компенсируют отсутствие движения тем, что сами синтезируют себе пищу, используя уникальные органеллы — хлоропласты — для превращения CO2 в соединения, которые подвергаются дальнейшему метаболизму или используются для получения полимеров клеточной стенки. Жесткость клеточных стенок обусловливает неподвижную форму существования растений, однако обеспечивает прочность их форм. Способ, каким клетки растения организуют и перестраивают свои стенки, определяет их форму. Это достигается за счет контроля направления роста.
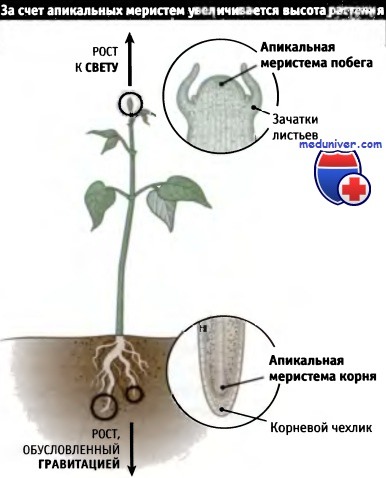
В отличие от животных, растения растут всю жизнь. Небольшие группы клеток, называемые меристемами, постоянно делятся и образуют новые клетки, необходимые для роста. Первичные меристемы на конце корня и побегов обеспечивают рост растения в высоту, а вторичная меристема боковой поверхности дает ему возможность увеличиваться в толщину. Меристема на конце побега постоянно образует модули, каждый из которых формируется выше предыдущего и состоит из листа или бутона и участка стебля, на котором они расположены. Этот тип роста зависит от внешних условий и позволяет растению на протяжении жизни изменять свою форму. Хотя деление клеток, благодаря которому растет растение, происходит в отдельных его частях, многие клетки сохраняют способность к делению при необходимости, в результате чего могут закладываться новые органы или даже совершенно новое растение.
Деление клеток, за счет которого происходит рост растений,
ограничивается специальными небольшими участками (меристемами),
которые расположены на кончиках корней и побегов.
Строение меристем показано на врезках.
Клетки первичной меристемы проходят через последовательные циклы процессов деления и роста, в результате чего образуются столбики или слои клеток расположенные вдоль оси направления роста растения. В отличие от клеток животных, жесткость клеточной стенки у растений не позволяет делиться с помощью образования перетяжки. Вместо этого поперек внутренней части клетки создается новая клеточная стенка — клеточная пластинка. Плоскость деления клетки определяется заранее препрофазным кольцом — состоящим из микротрубочек и актиновых филаментов кортекса. Эти компоненты образуются и исчезают до начала митоза, что свидетельствует об участии кортекса в этом процессе. Внутри клетки с крупными вакуолями, в поперечном направлении образуется слой цитоплазмы — фрагмосома. Это происходит одновременно с формированием препрофазного кольца, однако фрагмосома остается в клетке в течение всего митоза, позволяя формироваться митотическому веретену и клеточной пластинке.
В клетках растений, так же как и в клетках животных, в процессе деления формируется митотическое веретено. Однако у растений полюса веретена выглядят более размытыми из-за отсутствия в клетках центриолей и центросом. Когда в клетке завершается анафаза, между группами хроматид образуется фрагмопласт, который у растений представляет собой аппарат цитокинеза. Он состоит из двух рядов микротрубочек, ориентированных в противоположном направлении, причем концы их перекрываются в небольшой области. Везикулы, образующиеся в аппарате Гольджи, движутся вдоль микротрубочек и в этой области сливаются, образуя клеточную пластинку. Последняя растет в направлении от центра клетки и достигает ее стенки, сливаясь с ней. По мере роста клеточной пластинки, она пересекается трубочками ЭПР. При этом образуются каналы — плазмодесмы — посредством которых соединяется цитоплазма двух сестринских клеток и осуществляется обмен различными метаболитами.
По мере того как старые клетки апикальной меристемы прекращают делиться, они начинают растягиваться, причем часто достигают размера, во много раз превышающего исходный. В результате такого мощного расширения клеток, расположенных непосредственно за делящимися, точка роста продвигается все дальше в окружающую среду. Расширение зависит от поступления воды в клетки из-за разницы в осмотическом давлении. Избыток воды запасается в вакуолях — мембранных органеллах клеток растений. Вакуоли могут занимать около 95% всего клеточного объема. В результате набухания вакуолей в клетке развивается давление — тургор, которому противостоят прочные микрофибриллы клеточной стенки. Благодаря неслучайному характеру распределения целлюлозы тургорное давление преобразуется в усилие, направленное в определенную сторону. Микрофибриллы целлюлозы, расположенные перпендикулярно длинной оси клетки, обеспечивают «укрепляющую обвязку», которая предотвращает расширение клетки, но позволяет ей удлиняться и элонгировать. Целлюлоза синтезируется при участии мультиферментых комплексов, находящихся в плазматической мембране, и собирается в микрофибриллы. В самой клеточной стенке прочные целлюлозные микрофибриллы связаны между собой гликанами, и волокнистый компонент погружен в гель, образованный пектинами. Наряду с этими тремя типами полисахаридов, клетка содержит структурные белки и ферменты. Белок экспансии обеспечивает реформирование клеточной стенки, разрушая связи между соседними микрофибриллами и позволяя им расходиться.
Расположение целлюлозных микрофибрилл клеточной стенки часто определяется микротрубочками кортекса, связанными с внутренней поверхностью плазматической мембраны. Считается, что по ним движутся белковые комплексы синтеза целлюлозы, которые во время перемещения по поверхности мембраны образуют микрофибриллы. В отличие от многих клеток животных, в клетках растений микротрубочки не выходят из одной центральной точки, а образуются во множественных сайтах нуклеации, рассеянных по всему кортексу (и вокруг ядра). Такое диффузное расположение сайтов сборки интерфазных микротрубочек соответствует необходимости образования новой клеточной стенки при увеличении поверхности клетки. В кортексе микротрубочки располагаются параллельно друг другу и характеризуются высокой динамичностью, что позволяет им перегруппировываться. Обычно перегруппировка происходит при изменении направления расширения клеток. Она может быть вызвана изменениями освещенности и гравитации, подобно тому как это происходит под действием гормонов растений.
Секреторная система у растений также имеет децентрализованный характер. В отличие от клеток животных, для которых характерен единый аппарат Гольджи, в клетке растений их содержится очень много, и они рассеяны по всей клетке. Такой характер расположения аппарата Гольджи соответствует необходимости формирования новой плазматической мембраны и клеточной стенки на большом протяжении поверхности клетки при ее расширении. В клетке стопки Гольджи перемещаются по актиновым нитям. У растений актиновые филаменты расположены параллельно микротрубочкам, а также находятся в составе больших пучков, которые в клетке достигают значительной длины. Они обеспечивают постоянное и интенсивное движение везикул и органелл в ходе процесса, который называется ток цитоплазмы. Вероятно, это движение обеспечивается за счет миозина, находящегося на поверхности движущихся частиц.
Дифференцировка клеток происходит после их роста и иллюстрируется на примере образования клеток ксилемы. Программированная гибель слоев клеток, расположенных по центру оси роста, сопровождается утратой их поперечных стенок и формированием непрерывных трубок, по которым вода и питательные компоненты проходят от корней к надземным частям растения. Еще до гибели клеток микротрубочки кортекса объединяются в пучки, обеспечивающие образование утолщений клеточной стенки, которые укрепляют каждую трубку и не позволяют деформироваться ее стенкам. В отличие от первичных стенок, характерных для расширяющихся клеток, такие вторичные стенки у дифференцированных клеток состоят из дополнительных слоев целлюлозы и за счет их химической модификации лигнином обладают прочностью и водонепроницаемостью.
Хотя у растений большинство клеток растут за счет диффузного расширения, у некоторых из них, например у клеток корневых волосков и пыльцевых трубок, рост происходит на небольшом участке верхушечной точки. Этот участок элонгирует, образуя трубку, которая растет только на конце. По всей ее длине проходят микротрубочки и актиновые филаменты, и по последним к верхушечным точкам доставляются везикулы, обеспечивающие направленный рост плазматической мембраны.
В клетках растений содержатся уникальные органеллы, называемые пластидами. В процессе дифферен-цировки неспециализированные предшественники этих органелл — пропластиды — в зависимости от типа клеток образуют различные типы пластид. Такие пигментированные пластиды, как хромопласты, обеспечивают окраску цветов; в неокрашенных амилопластах запасается крахмал. Пластиды возникли в процессе эволюции за счет фагоцитоза клетками примитивных эукариот фотосинтезирующих прокариот. Прокариотическое происхождение пластид проявляется в том, что их белки и генетические элементы близко напоминают эти же компоненты клеток современных бактерий. К числу наиболее распространенных типов пластид относятся хлоропласта. Хлоропласта являются высокоспециализированными органеллами. Система их внутренних мембран (которые называются граны) отличается сложной внутренней структурой, которая обеспечивает начальные этапы процесса фотосинтеза.
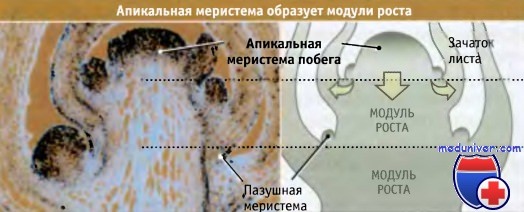
Фотография продольного среза растущего кончика побега антирринума (львиного зева).
Места интенсивного деления клеток выделены темным цветом.
Проводили гибридизацию in situ с циклиновой мРНК, в результате чего метились апикальная и пазушная меристема.
Диаграмма справа иллюстрирует, каким образом по мере роста растения апикальная меристема многократно образует модули роста.
Стрелки обозначают, что за счет деления клеток в центре апикальной меристемы в каждом модуле образуются клетки ствола,
в то время как при делении краевых клеток возникают зачатки, развивающиеся в листья или цветы.