Влияние кислорода на дыхательный центр. Роль кислорода в регуляции дыхания
а) Отсутствие влияния кислорода в регуляции деятельности дыхательного центра. Изменения в концентрации кислорода почти не имеют такого прямого влияния на дыхательный центр, который мог бы изменить пусковую импульсацию дыхания, хотя изменения концентрации кислорода оказывают на дыхательный центр непрямое влияние через периферические хеморецепторы.
Буферная система гемоглобин-кислород предоставляет тканям почти нормальное количество кислорода даже тогда, когда PO2 в легких поднимается от 60 до 1000 мм рт. ст. В связи с этим, несмотря на изменения легочной вентиляции в диапазоне от несколько ниже половины нормального объема до немного превышающего 20-кратный нормальный объем, возможно адекватное снабжение кислородом, исключая пребывание в специальных условиях. Для двуокиси углерода это не так, поскольку изменение вентиляции легких ведет к обратно пропорциональному изменению PCO2 и тканей, и крови, поэтому в процессе эволюции в качестве фактора регуляции дыхания была выбрана двуокись углерода, а не кислород.
Если ткани страдают от недостатка кислорода, имеется специальный механизм регуляции дыхания, расположенный в периферических хеморецепторах вне мозгового дыхательного центра. Этот механизм включается при слишком большом снижении количества кислорода в крови — обычно при снижении PO2 ниже 70 мм рт. ст..
б) Периферическая система хеморецепторов в регуляции дыхательной активности. Роль кислорода в регуляции дыхания. Кроме регуляции дыхательной активности самим дыхательным центром существует еще один механизм регуляции дыхания — периферическая система хеморецепторов (для облегчения понимания просим вас изучить рисунок ниже).
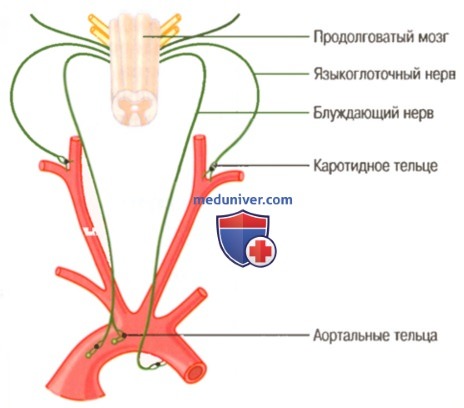
Влияние на дыхание расположенных в каротидных и аортальных тельцах периферических хеморецепторов
Специальные рецепторы для восприятия химических раздражений (хеморецепторы) расположены в нескольких зонах за пределами мозга. Они особенно важны для слежения за содержанием кислорода в крови, хотя с меньшей чувствительностью реагируют и на изменения в концентрации двуокиси углерода и ионов водорода. Сигналы от хеморецепторов передаются по нервам в расположенный в мозге дыхательный центр и помогают регуляции дыхательной активности.
Большинство хеморецепторов расположены в каротидных тельцах, немного хеморецепторов присутствуют и в аортальных тельцах (для облегчения понимания просим вас изучить рисунок выше), очень малое количество локализуются в других артериях грудной и брюшной частей тела.
Каротидные тельца расположены билатерально в бифуркациях общих сонных артерий. Их афферентные нервные волокна поступают через нервы Геринга в языкоглоточные нервы и далее — в дорсальную дыхательную зону продолговатого мозга. Аортальные тельца расположены вдоль дуги аорты, и афферентные нервные волокна от них проходят в составе блуждающих нервов также в дорсальную дыхательную зону продолговатого мозга.
Каждое хеморецептивное тельце снабжается кровью через маленькую артерию, отходящую непосредственно от близлежащего артериального ствола. Кровоток в этих артериях исключительно интенсивен, и масса протекающей крови за 1 мин превышает собственную массу тельца в 20 раз, поэтому количество кислорода в протекающей крови практически не снижается и хеморецепторы соприкасаются фактически только с артериальной, а не венозной кровью, и PO2 в них равно PO2 артериальной крови.
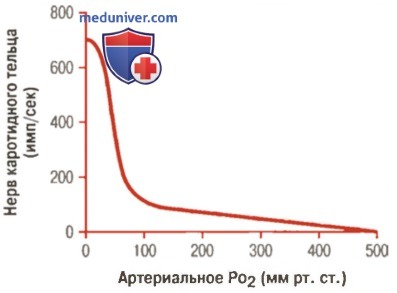
Влияние PO2 в артериальной крови на количество импульсов от каротидного тельца у кошки
в) Стимуляция хеморецепторов снижением количества артериального кислорода. Падение концентрации кислорода в артериальной крови ниже нормы сильно стимулирует хеморецепторы. Это хорошо видно на рисунке выше, демонстрирующем влияние разных величин артериального PO2 на количество импульсов, передаваемых от каротидного тельца. Обратите внимание, что количество импульсов особенно чувствительно к изменениям артериального PO2 в пределах 60-30 мм рт. ст., т.е. в диапазоне, где насыщение гемоглобина кислородом стремительно падает.
г) Влияние концентраций двуокиси углерода и ионов водорода на активность хеморецепторов. Повышение концентрации как двуокиси углерода, так и ионов водорода также раздражает хеморецепторы и этим повышает дыхательную активность непрямым способом. Однако прямое воздействие этих факторов на дыхательный центр намного сильнее (примерно в 7 раз), чем опосредованное хеморецепторами, поэтому в практике непрямое влияние двуокиси углерода и ионов водорода с участием хеморецепторов можно не учитывать. Имеется одно различие между периферическим и центральным влиянием двуокиси углерода: стимуляция периферических хеморецепторов происходит в 5 раз быстрее, чем центральная стимуляция, поэтому периферические хеморецепторы имеют важное значение для повышения скорости ответной реакции на двуокись углерода в начале физической работы.
д) Основной механизм стимуляции хеморецепторов дефицитом кислорода. Точный механизм, при помощи которого низкое PO2 раздражает нервные окончания в каротидных и аортальных тельцах, до сих пор неизвестен. В этих тельцах находится множество железистых клеток (glomus cells) характерного вида, которые имеют прямую или непрямую синаптическую связь с нервными окончаниями. Некоторые исследователи полагают, что эти клетки могут работать хеморецепторами и стимулировать нервные окончания. В других исследованиях было показано, что сами нервные окончания обладают прямой чувствительностью к понижению PO2.
Видео физиология регуляции дыхания - профессор, д.м.н. П.Е. Умрюхин