У новорожденных, как и у взрослых, в почках должны реабсорбироваться весь отфильтровавшийся бикарбонат и экскретироваться все образующиеся в процессе метаболизма кислоты (у взрослых — примерно 1 мэкв/кг/сут). У детей образование кислот повышено вследствие более интенсивного метаболизма и роста костей. У новорожденных сывороточная концентрация бикарбоната составляет примерно 19 мэкв/л, а у недоношенных может быть снижена до 15 мэкв/л.
Такая низкая концентрация обусловлена малой максимальной скоростью канальцевой реабсорбции бикарбоната. Из-за этого новорожденные предрасположены к развитию тяжелого метаболического ацидоза при больших потерях бикарбоната (например, при поносе).
У взрослых проксимальная канальцевая секреция Н+ на 2/3 осуществляется Na+/Н+-обменником апикальной мембраны и на 1/3 — Н+-АТФазой апикальной мембраны. Секретируемый Н+ связывается с бикарбонатом в просвете канальца с образованием Н2СO3. Карбоангидраза апикальной мембраны катализирует распад Н2СO3 до СO2 и Н2O. СO2 диффундирует в клетки проксимальных канальцев, где под действием карбоангидразы цитоплазмы превращается в Н2СO3 и затем — в бикарбонат.
Большинство отфильтровавшегося бикарбоната реабсорбируется в проксимальных канальцах. Скорость этой реабсорбции у новорожденных примерно в 3 раза ниже, чем у взрослых. Это объясняется тем, что у новорожденных низка активность и Nа+/Н+-обменника, и Н+-АТФазы. Скорость котранспорта Na+-НСО3 через базолатеральную мембрану относительно высока и потому не ограничивает реабсорбцию бикарбоната.
Активность карбоангидразы по мере роста возрастает, однако неизвестно, ограничивает ли низкая активность этого фермента скорость реабсорбции бикарбоната у детей. Как бы то ни было, низкая скорость реабсорбции бикарбонатов в проксимальных канальцах является основным фактором, обусловливающим низкий почечный порог для бикарбоната у новорожденных.
У взрослых в зависимости от КНДР в корковом отделе собирательной трубочки бикарбонаты могут реабсорбироваться или секретироваться. Секреция Н+ осуществляется вставочными клетками типа А, а секреция бикарбонатов — вставочными клетками типа В, в апикальной мембране которых имеется Cl/HCO3-обменник, а в базолатеральной мембране — Н+-АТФаза. По мере созревания количество вставочных клеток обоих типов увеличивается.
Корковый отдел собирательной трубочки переходит в мозговой отдел, в котором также имеются вставочные клетки. Число этих клеток и скорость секреции Н+ в этом отделе у взрослых и новорожденных примерно одинаковы.
Выведение кислот, образующихся у детей в результате метаболизма и роста костей, зависит от способности к секреции Н+ в дистальном отделе нефрона. У новорожденных КЩР частично поддерживается за счет преимущественного потребления щелочных продуктов. Секрети-руемые в дистальных канальцах Н+ связываются аммиаком и анионами титруемых кислот (например, фосфатом), играющими роль буферов.
У доношенных и недоношенных новорожденных в течение первых 7 сут после рождения способность к выведению кислот уменьшена из-за низкой скорости экскреции как титруемых кислот, так и аммиака. В проксимальных канальцах у них замедлено образование аммиака из глутаминовой кислоты и в результате снижена доставка аммиака в дистальный отдел нефрона. У вскармливаемых грудью новорожденных экскреция титруемых кислот ограниченна еще и потому, что содержание фосфата в женском молоке ниже, чем в коровьем. Наконец, у новорожденных, в отличие от взрослых, выведение кислот находится почти на предельном уровне, и в результате у них отсутствует возможность увеличения экскреции аммиака и титруемых кислот (и, следовательно, Н+) в ответ на ацидоз.
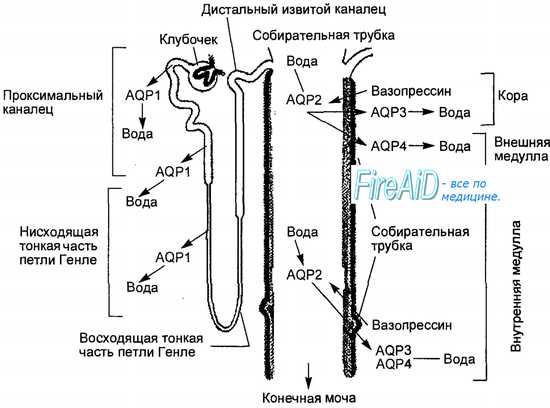
Локализация в нефроне и участие в транспорте воды через эпителий почечных канальцев аквапоринов (водных каналов) разных типов (AQP 1—4).
AQP1 — водные каналы, постоянно локализованные в проксимальных извитых канальцах и нисходящем отделе петли Генле, обеспечивают трансэпителиальный транспорт воды из просвета канальцев, т. е. проксимальную реабсорбцию воды.
AQP2— вазопрессинозависимый тип аквапоринов. Эти водные каналы встраиваются в люминальную мембрану главных клеток собирательных трубочек только при наличии вазопрессина, обеспечивая зависимую от вазопрессина дистальную реабсорбцию воды и концентрирование мочи.
AQP3 и AQP4— водные каналы, постоянно локализованные в базолатеральных мембранах главных клеток эпителия собирательных трубочек, обеспечивают транспорт воды из эпителиальных клеток в интерстиций мозгового вещества.