Модификация и сворачивание белков после трансляции
Котрансляционные и посттрансляционные модификации представляют собой важные этапы транслокации, активации, регуляции и распада белков. В действительности существует, вероятно, лишь небольшое количество белков, которые не подвергаются каким-либо химическим изменениям во время синтеза или по его завершении. Из многих белков удаляется N-концевой остаток метионина, а прочие излишние участки некоторых белков подвергаются расщеплению.
Другие распространенные модификации включают (но не ограничиваются ими) гликозилирование, ацетилирование, жировое ацилирование и формирование дисульфидных связей. Несмотря на повсеместное присутствие модификации белков в природе, многое из того, каким образом эти изменения влияют на деятельность каждого отдельно взятого белка, остается неизвестным.
Это объясняется главным образом тем, что единичный вид модификации запускает различные функции в зависимости от целевого белка. Например, аспарагин-связанное гликозилирование может стабилизировать процесс сворачивания одного полипептида, тогда как у другого белка оно может играть важную роль в распознавании клетки или ее функции.
Механизм этих и других изменений белка в процессе трансляции и после нее разнообразен и сложен. Однако важно отметить, что мутации в генах, участвующие в модификациях белков, приводят к разнообразным расстройствам, например врожденным нарушениям гликозилирования типа I, мультисистемному заболеванию, при котором происходит аномальное гликозилирование гликопротеинов слизистой оболочки кишечника.
Нарушения гликозилирования типа I проявляются такими кишечными симптомами, как воспаление и аномальный транспорт липидов энтероцитами или нарушение кишечной проницаемости. Эти заболевания у человека подчеркивают тот факт, что котрансляционные и посттрансляционные модификации являются тесно связанными событиями в регулировании синтеза белка, а также его распада.
Трансляции мРНК в основном начинаются на цитозольных рибосомах. В отдельных классах белков образующийся полипептид нацелен (с помощью распознающей сигнал частицы) на эндоплазматический ретикулум для завершения обработки и осуществления некоторых трансляционных и посттрансляционных модификаций. К этим классам белков относятся белки ЭПР и резидентные белки аппарата Гольджи, белки, связанные с мембраной, белки эндосомнолизосомальной системы и белки, предназначенные для секреции.
После трансляции и модификации образующиеся пептиды, расположенные в ЭПР, должны быть правильно свернуты и собраны до того, как перейдут во внутриклеточные органеллы либо на поверхность клетки. Семейство белков, известное как молекулярные шапероны (chaperones), решает эту задачу, функционируя как система «контроля качества» на стадии образования пептидов.
Несвернутые или неправильно свернутые белки на время задерживаются в просвете эндоплазматического ретикулума (ЭПР) в ожидании правильной компоновки. Белки, которые не могут быть свернуты правильно, перемещаются из ЭПР для целенаправленной деградации в цитозоле протеосомами.
Для гомеостаза ЭПР необходимо, чтобы емкость аппарата сворачивания белка оставалась в состоянии баланса с потребностями. Этот баланс может быть нарушен в результате стресса, вызванного состоянием окружающей среды (например, недостатком глюкозы или нарушением гомеостаза Са2+ во время гипоксии), генетическими мутациями в белках, которым необходима сборка в ЭПР (например, CFTR), или нарушениями в самом аппарате сворачивания белка.
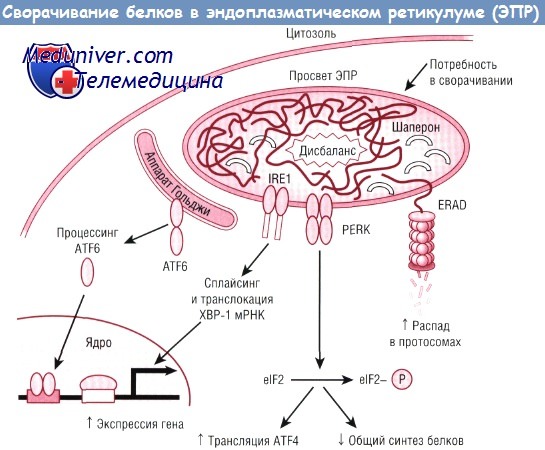
Дисбаланс между потребностью в сворачивании и сворачивающей способностью создает в эндоплазматическом ретикулуме (ЭПР) стресс, приводящий у млекопитающих к тройственной реакции несвернутых белков (стрессовая реакция ЭПР).
Триада стрессовых рецепторов отслеживает изменения, происходящие в ЭПР и устанавливает временные модели экспрессии генов, которые совместными усилиями активируют обработку, накопление и транспорт секреторных белков, несмотря на подавление синтеза новых белков.
Один из стрессовых сенсоров, трансмембранный фактор транскрипции (ATF) 6, представляет собой связанный с мембраной аппарата Гольджи белок, который после протеолитического расщепления направляется в ядро, чтобы управлять транскрипцией генов, подвергающихся UPR.
Вторым сенсором является IRE1, трансмембранная протеинкиназа ЭПР, РНКазная активность которой важнее для сплайсинга ХВР-1 мРНК в мощный фактор транскрипции, который также руководит генами, нацеленными на UPR.
Третьим сенсором UPR является PERK, другая трансмембранная протеинкиназа ЭПР, которая угнетает общий синтез белков путем фосфорилирования elF2. Возникающий в результате этого пониженный синтез белков будет предупреждать дальнейшую перегрузку ЭПР и давать клетке достаточно времени для того, чтобы перестроить экспрессию генов.
elF — эукариотический фактор инициации; ERAD — расщепление белка, связанное с эндоплазматическим ретикулумом; IRE1 — трансмембранный белок, выполняющий функции и киназы, и эндонуклеазы; PERK — PKR-подобная протеинкиназа эндоплазматического ретикулума; ХВР-1 — X-box-связывающий белок 1; мРНК — матричная РНК.
Когда этот баланс нарушается, несвернутые или неправильно свернутые белки накапливаются на мембране эндоплазматического ретикулума (ЭПР) как «клеточный мусор». В результате инициируется адаптивный механизм, который называют реакцией несвернутых белков (UPR) или стрессовой реакцией ЭПР. UPR включает три отдельных процесса.
Во-первых, снижается общая скорость синтеза белка для уменьшения нагрузки на эндоплазматический ретикулум (ЭПР) по сворачиванию белков и непосредственного ингибирования трансляции циклина D1, способствуя тем самым приостановке клеточного цикла.
Во-вторых, шапероны, резиденты эндоплазматического ретикулума (ЭПР), транскрипционно индуцированы для увеличения сворачивающего потенциала в этих органеллах.
В-третьих, для удаления накапливающихся клеточных отходов инициируется система расщепления белка, связанного с эндоплазматическим ретикулумом. Активация ERAD имеет важное значение для распада несвернутых белков через убиквитин-протеасомный путь. Если стресс ЭПР не может быть уменьшен этими тремя процессами, тогда активируется клеточный механизм, вызывающий апоптоз.
Накопление несвернутых белков распознают несколько резидентных молекул эндоплазматического ретикулума (ЭПР). Одной из них является еIР2-киназа PERK. Как было упомянуто ранее, PERK индуцирует фосфорилирование фактора трансляции eIF2, что приводит к выключению трансляции на начальных этапах. Эта реакция блокирует синтез новых белков и предупреждает дальнейшее накопление несвернутых белков, образующих токсические скопления.
В это же время увеличение фосфорилирования eIF2 делает возможной трансляцию специфической мРНК, которая кодирует ATF4. ATF4 способствует транскрипции генов, уменьшающих стресс, а также генов, которые связаны с апоптозом.
Стимуляция трансляции специфической мРНК вопреки общей репрессии является скоординированным процессом (например, общей стрессовой реакцией), что подчеркивает способность клетки в организованном порядке разделять и направлять в определенное русло множественные биологические процессы. Кроме того, на накопление несвернутых белков реагируют и другие резидентные белки ЭПР, такие как IRE1 (трансмембранный белок, выполняющий функции и киназы, и эндонуклеазы) и ATF6, путем активации транскрипции других генов, и все это увеличивает сворачивающий потенциал в ЭПР.
Хотя клеточный стресс, вне сомнения, является основным активатором UPR, следует отметить, что UPR вызывается не только стрессовыми событиями. Напротив, последние исследования показывают, что дифференцировка клеток, связанных с увеличением производства секреторных белков, сочетается с активацией тех же самых сигнальных путей. Например, для конечной дифференцировки В-лимфоцитов в зрелые плазматические клетки, которые секретируют антитела, необходимо, чтобы ЭПР набирал и выделял большое количество антител с исключительной эффективностью.
При дифференцировке В-клеток нужно огромное увеличение белоксворачивающей способности ЭПР. При переходе В-клеток к секреции антител с высокой скоростью UPR регулирует гомеостаз ЭПР и события, необходимые для гуморального иммунитета.
Ясно также, что UPR необходима для нормальной функции клеток. Пациенты с синдромом Уолкотта-Раллисона с периода новорожденности и раннего детства страдают тяжелой формой инсулинозависимого диабета, а также имеют дефекты в формировании костей и замедление темпов роста. Генетическая мутация у этих пациентов затрагивает ген EIF2AK3, который у человека кодирует еIF2-киназу PERK.
У мышей с функциональным нарушением гена EIF2AK3/PERK выявляется массовый апоптоз экзокринной части поджелудочной железы спустя несколько недель после рождения, приводящий к уничтожению производящих инсулин b-клеток.
Некоторые исследования показывают, что сигнальные пути, связанные с UPR, эволюционировали, чтобы соответствовать потребностям в питании при некоторых метаболических процессах. Клеточная реакция на стресс, которая связана с реакцией несвернутого белка, но отличается от нее, включает реакцию клеток на недостаток аминокислот.
В ответ на ограничение поступления незаменимых аминокислот еIF2-киназа GCN2 фосфорилирует фактор трансляции, чтобы снизить уровень синтеза белков и инициировать каскад событий, не связанных со сворачиванием белка, что приводит к повышению экспрессии генов, участвующих в метаболизме и транспорте аминокислот. Животные с отсутствием GCN2 не в состоянии справиться с нехваткой незаменимых аминокислот в рационе и, находясь на диете, лишенной лейцина, через несколько дней начинают агонизировать. Таким образом, лишение аминокислоты или голодание активирует сигнальный путь клеточного стресса, который включает компоненты UPR.
В дополнение к активации фосфорилирования eIF2 недостаток аминокислот вызывает передачу каскада сигналов на mTOR, чтобы снизить его чувствительность к инсулину и факторам роста. Доминирующим проксимальным регулятором сигналов для TORC1 и активности киназы является ген Rheb (ras-like small GTPase).
Истощение аминокислотного запаса препятствует способности Rheb активировать mTOR, что действенно ингибирует рост сигналов, вызванных последующими звеньями сигнального пути Rheb, а именно передачу сигнала, связанную с инсулиновым путем.
Стимулирование mTOR сигналами для содействия увеличению анаболизма через биогенез рибосом, синтез белков, импорт нутриентов и ускорение клеточного цикла требует удвоенного поступления как инсулина и факторов роста, так и аминокислот. Поэтому важным моментом в питании новорожденных является то, что для нормального функционирования основных механизмов роста человека как in vitro, так и in vivo чрезвычайно важны экзогенные аминокислоты.