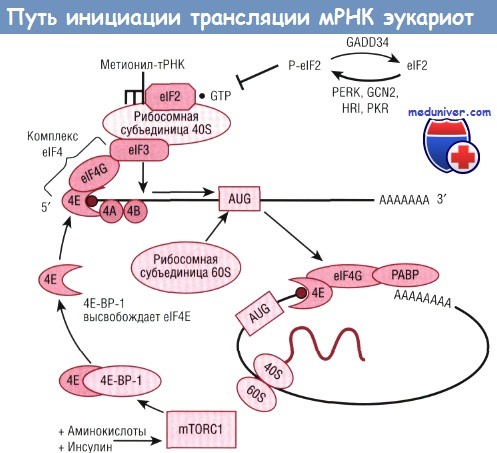
Большинство факторов контроля трансляции относятся к стадии инициации, которая, в свою очередь, может быть разделена на три этапа, определящих всю деятельность стадии инициации. Первый этап обусловлен связыванием инициирующей тРНК (в частности, метионил-тРНК) и малых рибосомных субъединиц. Метионил-тРНК доставляется к рибосомной субъединице 40S белковым фактором eIF2.
Способность eIF2 и гуанозин-5'-трифосфата (GTP) соотноситься с инициирующей метионил-тРНК регулируется состоянием фосфорилирования eIF2. Прирост фосфорилированных eIF2 предотвращает совмещение метионил-тРНК с небольшой рибосомной субъединицей, что приводит к общему подавлению трансляции мРНК. В условиях клеточного стресса фосфорилирование eIF2 также сигнализирует о необходимости увеличения экспрессии конкретных мРНК, которые работают, чтобы регулировать и смягчать клеточный стресс.
Эти процессы относятся к комплексной реакции на стресс и инициируются семейством четырех еIF2-киназ: PERK (PKR-подобной протеинкиназой эндоплазматического ретикулума, также называемой pancreatic eIF2 kinase, РЕК), которая распознает стресс эндоплазматического ретикулума (ЭПР); GCN2 (general control nonderepressible-2 kinase), которая активируется недостатком аминокислот и ультрафиолетовым излучением; гем-контролируемым ингибитором, который обнаруживает утрату тема; PKR (дцРНК-зависимой протеинкиназой), которая активируется вирусной инфекцией.
Эти протеинкиназы функционируют в первую очередь для распознавания конкретных стрессоров окружающей среды, но указанные протеины, как обнаружили, действуют совместно и имеют возможность осуществлять вспомогательную деятельность киназы, когда функционирование первичной реагирующей киназы нарушено или отсутствует.
Второй этап в инициации трансляции, который является объектом регулирования, заключается в присоединении малой рибосомной субъединицы к выбранной мРНК. Для этого необходим многокомпонентный комплекс, имеющий общее название eIF4 (или eIF4F). Один из белков в этой группе, называемый eIF4E, выбирает мРНК, чтобы быть транслированным путем связывания ее 5'-кэп-структуры. Все эукариотические мРНК имеют кэп-структуры 7-метилгуанозина.
Эукариотический фактор инициации (elF) 2, связанный с гуанозин-5'-трифосфатом (СТР), необходим для присоединения его и инициирующующей тРНК (метионил-тРНК) к рибосомной субъединице 40S. Это событие регулируется фосфорилированием elF2, осуществляемым семейством протеинкиназ, которые ингибируют комплекс перед инициацией в условиях клеточного стресса.
После формирования комплекса перед инициацией намеченная для трансляции мРНК отбирается группой факторов трансляции elF4, которая включает мРНК-кэпсвязывающий белок, elF4E и «каркас» белка, elF4C. elF4C играет важную роль в процессе тесного сближения малых субьединиц рибосомы и мРНК путем взаимодействия с elF4E и elF3, а также в циркуляризации мРНК посредством связывания поли-(А)-связывающего белка (РАВР).
Нутриент-зависимый комплекс 1 mTOR (mTORCl) регулирует активность этого этапа частично путем регулирования связи elF4C с elF4E по отношению к репрессору трансляции 4Е-ВР-1. Вслед за успешным формированием активного комплекса elF4 и выбором мРНК рибосомная субъединица 60S присоединяет субъединицу 40S и образует компетентную рибосому 80S. HRI — гем-контролируемый ингибитор; PERK — PKR-подобная протеинкиназа эндоплазматического ретикулума; PKR — дцРНК-зависимая протеинкиназа.
Ковалентно присоединенная молекула 7-метилгуанозина служит для защиты мРНК от экзонуклеаз, но, что более важно, 7-метилгуанозин распознается мРНК-кэпсвязывающим белком (eIF4E), чтобы осуществить отбор и связывание с малой рибосомной субъединицей. Второй член группы eIF4, называемый eIF4G, способствует сближению малой рибосомной субъединицы и мРНК. Он решает эту задачу, выступая в качестве «каркаса» (scaffold), состоящего из 13 субъединиц белкового комплекса, присоединяющего eIF4E, eIF4A и eIF3, тем самым облегчая их связи с рибосомой 40S. Семейство репрессорных белков (4Е-ВР, в частности 4Е-ВР-1) может предотвратить взаимодействие eIF4G и eIF4E и тем самым затормозить связывание рибосомы 40S и мРНК.
Второй функцией eIF4G является соединение с поли-(А)-связывающим белком, т.е. белком, который связывает участок из 20-250 остатков аденозина на 3'-конце мРНК. Эти два связанных события завершаются 5' => 3'-циркуляризацией мРНК в процессе трансляции. Считается, что циркуляризация мРНК важна для стабилизирования рекрутированных рибосомных субъединиц 40S, а также в целях эффективной утилизации рибосом на стадии терминации для очередного цикла трансляции с использованием той же мРНК. Таким образом, взаимодействие eIF4G с поли-(А)-связывающим белком вовлечено как в усиление образования инициированных комплексов 40S и 80S, так и в рециркуляцию рибосом путем циркуляции мРНК.
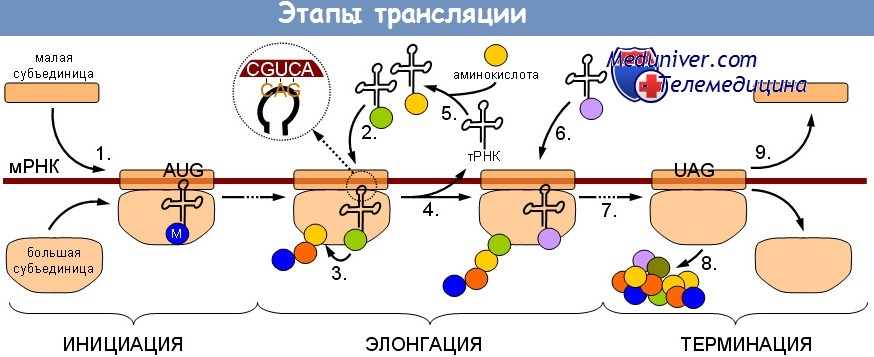
Заключительный этап стадии инициации трансляции предполагает объединение малых рибосомных субъединиц (связанных с мРНК) с большой рибосомной субъединицей. Это событие катализируется несколькими eIF, в том числе eIF2, eIF3 и eIF5. Эти факторы и их взаимодействия были изучены, но детали, непосредственно касающиеся возможного регулирования на этой стадии трансляции, выходят за рамки данного издания.
В отношении кормления новорожденных следует сказать, что для стимулирования синтеза белка на уровне инициации трансляции необходимы как инсулин, так и аминокислоты. Аминокислоты, в частности лейцин (аминокислота с разветвленной цепью), служат в качестве сигнальных молекул, которые регулируют начальные этапы трансляции мРНК, направляя образование комплексов eIF4 и фосфорилирование eIF2.
Единственное, что нужно для стимуляции образования eIF4 и синтеза белка в скелетных мышцах новорожденных поросят, — это физиологическое повышение уровня лейцина. Данный вывод является крайне важным для организации питания новорожденных, поскольку он означает, что аминокислоты в клетке имеют чрезвычайно высокое значение для стимуляции роста истощенных тканей. Каким именно образом аминокислоты инициируют сигнал для включения трансляционного аппарата, до сих пор остается загадкой. Известно, что mTOR-киназа является одним из основных участников, выявляющих наличие в клетке аминокислот, и что стимуляция синтеза белка за счет обеспечения новорожденных или детенышей животных лейцином оптимальным образом активируется, когда сигнал аминокислоты сочетается с активацией инсулинового пути трансдукции сигнала.
Сигнальные пути, активированные инсулином и аминокислотами, сходятся на уровне mTOR, которая затем катализирует фосфорилирование 4Е-ВР-1. Фосфорилирование вызывает высвобождение 4Е-ВР-1 из связи с eIF4E, что позволяет eIF4G связать eIF4E и eIF3, образуя комплекс eIF4F. Эти процессы связывания частично или полностью блокируются путем обработки рапамицином, ингибитором mTOR. Стимулирование активности mTOR также приводит к активации фосфорилирования S6K1, что активирует биогенез рибосом и других процессов, которые могут приводить к увеличению размеров клеток. Этот сигнальный путь S6K1, реагирующий на аминокислоты или инсулин, полностью ингибируется рапамицином, тогда как связывающая активность eIF4G подавляется в меньшей степени.
Ясно, что организм новорожденного уже с момента рождения готов реагировать на поступление нутриентов активным и эффективным белковым синтезом. Однако компоненты трансляции и факторы, необходимые для активации роста анаболических процессов, с возрастом ослабевают. Такое снижение трансляционной способности в ответ на кормление у новорожденных детей происходит очень быстро (у новорожденных поросят — в течение нескольких дней). Эти данные подтверждают наблюдение о решающем значении раннего белкового питания, обеспечивая реальную возможность максимально стимулировать развитие организма у детей, родившихся с низкой массой тела.
Возможно, именно в силу этого обстоятельства недостаток белков в пище ребенка на раннем этапе жизни трудно преодолевается в дальнейшем, когда даже усиленное питание и заместительная гормональная терапия бывают не в состоянии устранить отставание в росте, т.к. способность организма реагировать на качество питания в процессе дальнейшей жизни существенно уменьшается.