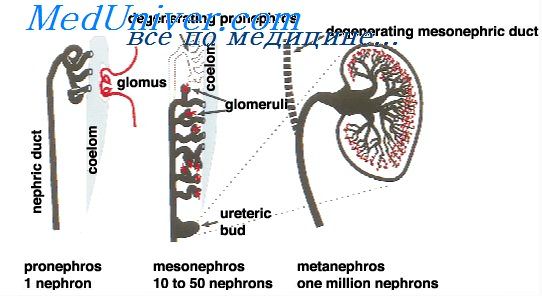
Дифференцировкой клеток и образованием трехмерной структуры почки при нефрогенезе управляет сложная система двусторонних сигналов между дифференцирующимися клетками и их окружением.
Глиальный нейротрофический фактор (GDNF). Взаимодействие этого фактора с рецепторным комплексом, состоящим из белков Ret и GDNFRa, — главный регулятор ветвления мочеточникового выроста вглубь мезенхимы метанефрогенной ткани. Экспрессия гена GDNF в мезенхиме контролирует рост мочеточникового выроста из вольфова протока, а кроме того, отвечает за ветвление выроста внутри мезенхимы. При направленной делеции гена GDNFy животных развиваются канальцы первичной почки и нормально формируется вольфов проток, однако мочеточниковый вырост отсутствует. Без индукции со стороны мочеточникового выроста метанефрогенная ткань постепенно подвергается апоптозу, что ведет к развитию дефектной почки или ее агенезии.
Семейство генов PAX. Факторы транскрипции, кодируемые семейством генов PAX, имеют ДНК-связывающий домен, который называется парный бокс. Показано, что несколько генов семейства PAX важны при развитии почек у мышей и человека. Два из них, РАХ2 и РАХ8, экспрессируются в развивающейся почке с некоторым перекрытием во времени. РАХ2 экспрессируется в вольфовом протоке, мочеточниковом выросте и метанефрогенной ткани. Сходно с ним распределение РАХ8, но он начинает экспрессироваться немного позже, в образующихся при конденсации мезенхимы везикулах и S-образном клубочке.
Мутации гена РАХ2 у человека приводят к недоразвитию почек. У этого гена неполное доминирование: даже у гетерозигот его мутации могут проявляться в фенотипе. У человека описан синдром с аутосомно-доминантным наследованием, вызванный мутацией гена РАХ2 и включающий дисплазию почек, пузырно-мочеточниковый рефлюкс и колобому, причина которого — мутация в гене РАХ2.
Фактор транскрипции WT1. Ген-супрессор опухоли Вильмса (WT1) кодирует фактор транскрипции с ДНК-связывающим доменом типа «цинковый палец»; с его помощью мезенхима стимулирует отпочковывание мочеточникового выроста от вольфова протока. Этот ген начинает экспрессироваться в мезенхиме при ее конденсации рядом с клетками мочеточникового выроста, рост которых она стимулирует, но не в нем самом и продолжает экспрессироваться в формирующемся S-образном почечном клубочке.
WT1 подавляет экспрессию нескольких факторов роста, в том числе ИФР-II и А-цепи тромбоцитарного фактора роста. В начале конденсации экспрессия гена WT1 возрастает, благодаря чему нет избыточной пролиферации мезенхимных клеток. В отсутствие фактора WT1 метанефрогенная ткань могла бы расти безостановочно. Мутации гена WT1 у человека вызывают опухоль Вильмса (нефробластому), синдром Дени—Дрэша и синдром Фрайзера.
Белки семейства WNT. Эти белки стимулируют трансформацию метанефрогенной ткани в клетки эпителия. В растущем вольфовом протоке экспрессируется ген WNT11, но затем его экспрессия сосредоточивается в участке протока непосредственно рядом с метанефрогенной тканью. При появлении мочеточникового выроста ген WNT11 экспрессируется лишь на его конце, а затем всегда обнаруживается на кончиках растущих разветвлений. Ген WNT4 необходим для трансформации индуцированной мезенхимы в эпителий.
Гены ЕМХ2 и BF2. Ген ЕМХ2, содержащий гомеобокс, необходим для ветвления мочеточникового выроста. У мышей с мутацией этого гена мочеточниковый вырост удлиняется и взаимодействует с мезенхимой, но не способен ветвиться. Ген BF2 вначале экспрессируется в клетках мезенхимы, рядом с клетками, экспрессирующими ген РАХ2. Клетки, экспрессирующие BF2, становятся потом интерстициальными клетками, а клетки, экспрессирующие РАХ2, — клетками почечного эпителия. У мышей с мутацией гена BF2 на всем протяжении процесса образования коркового и мозгового вещества в нем продолжают сохраняться интерстициальные клетки. У таких мышей сильно подавлен процесс ветвления мочеточникового выроста.
Молекулы адгезии. Молекулы адгезии — это гликопротеиды, участвующие в межклеточных контактах; некоторые из них, такие как Е-кадгерин и NCAM (молекула адгезии нервных клеток), экспрессируются в почке. Взаимодействия между молекулами адгезии и гликопротеидами внеклеточного матрикса — важнейшие события в развитии почки, регулируемые интегринами; по-видимому, эти взаимодействия различны на разных стадиях нефрогенеза и в разных сегментах. Важную для развития почки роль могут играть также ламинины, протеогликаны и ферменты деградации внеклеточного матрикса.
Интегрины — главное семейство рецепторов к внеклеточному матриксу; это гетеродимерные белки, каждый из которых образован двумя цепями: а и b. Для развития почки важны а3b1- и а8b1-интегрины. В отсутствие а8b1-интегрина мочеточниковый вырост формируется, но не растет в направлении метанефрогенной ткани и не ветвится. В мочеточниковом выросте, собирательных трубочках, в эндотелии клубочков и подоцитах экспрессируется а3b1-интегрин. Результаты, полученные на мышах с направленной делецией гена а3b1 -интегрина, указывают на его роль в развитии подоцитов, формировании базальной мембраны клубочков и ветвлении мочеточникового выроста.
Факторы роста. Почкой эмбриона вырабатываются морфогены, влияющие на выживаемость клеток, их пролиферацию, дифференцировку и морфогенез, — факторы роста и цитокины. Список возможных регуляторов — в их числе многие секретируемые факторы и их рецепторы, либо экспрессирующиеся в эмбриональной почке, либо in vitro способные влиять на ее развитие, — продолжает удлиняться.
При блокировании ИФР-I наблюдается выраженная атрофия окончательной почки. In vitro воздействие ИФР-I вызывает трофический ответ, а исследования in vivo показали, что ИФР-I устраняет нарушения развития почек, вызванные подавлением ренин-ангиотензиновой системы. Эпидермальный фактор роста — белок, который вырабатывается главным образом слюнными железами и почками.
Он связывается с рецептором с собственной тирозинкиназной активностью, который связывает также трансформирующий фактор роста а — возможно, являющийся эмбриональной формой эпидермального фактора роста. Уровень экспрессии эпидермального фактора роста высок в постнатальном периоде, а уровень экспрессии трансформирующего фактора роста а выше в эмбриональном. При добавлении эпидермального фактора роста к органной культуре эмбриональной почки наблюдается гипертрофия почки и образуются кисты, но образование клубочков подавляется. Антитела к трансформирующему фактору роста а вызывают нарушения ветвления мочеточникового выроста и плохой рост почечной ткани.
Суперсемейство трансформирующего фактора роста b содержит большое число бифункциональных регуляторных белков, таких как трансформирующие факторы роста р,_5, белки — регуляторы морфогенеза кости, глиальный нейротрофический фактор (GDNF) и другие активаторы и ингибиторы роста. Добавление трансформирующего фактора роста р к культуре эмбриональной почечной ткани подавляет ее рост и образование клубочков и останавливает развитие проксимальных канальцев и собирательных трубочек. У мышей с делецией гена ВМР7 наблюдаются недоразвитие либо агенезия почек, глаз и костей. Эпидермальный фактор роста, фактор роста гепатоцитов, факторы роста фибробластов и трансформирующие факторы роста, а также их рецепторы участвуют в ветвлении мочеточникового выроста и образовании канальцев.
Апоптоз при нефрогенезе - развитии почек
Апоптоз важен для нормального развития почек. Большая часть клеток мезенхимы, если они не трансформировались в эпителий, подвергаются апоптозу.
Некоторые факторы роста способны защитить клетки метанефрогенной ткани от апоптоза, среди них эпителиальный фактор роста и основный фактор роста фибробластов. Есть онкогены, регулирующие рост путем влияния на апоптоз. К примеру, фактор транскрипции WT1 связывается с геном ТР53, ингибируя вызванный им апоптоз. У мышей, лишенных гена WT1, наблюдается апоптоз клеток мезенхимы. Активирующие мутации гена ТР53 ведут к олигонефронии и апоптозу.
В конденсирующейся мезенхиме и мочеточниковом выросте экспрессируется еще один протоонкоген — BCL2, а в дифференцированных эпителиальных клетках его экспрессия подавлена. У мышей, лишенных гена BCL2, наблюдаются недоразвитые кистозные почки и обширный апоптоз.
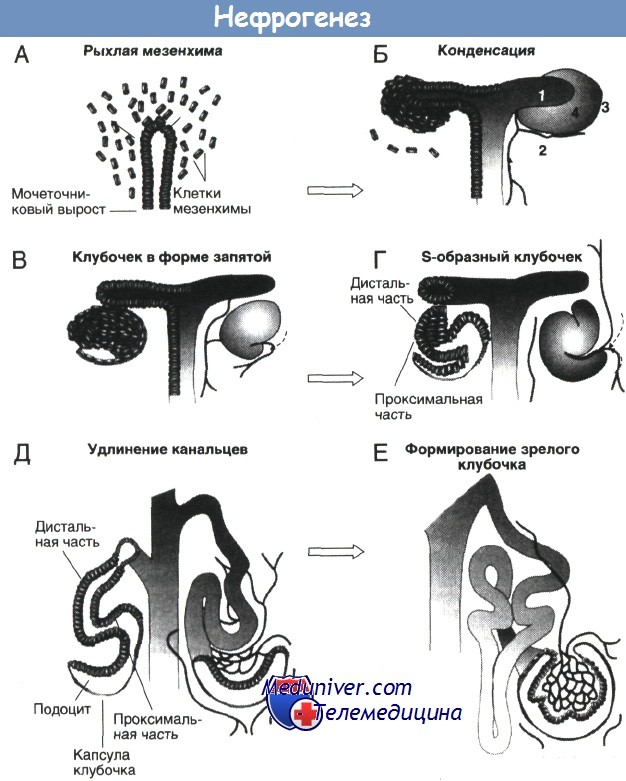
А, Б. Взаимодействие эпителия ветвящегося мочеточникового выроста с рыхлой мезенхимой метанефрогенной ткани приводит к конденсации мезенхимы. Цифрами на Б обозначены: 1 — эпителий мочеточникового выроста; 2 — кровеносные сосуды; 3 — недифференцированная мезенхима; 4 — конденсированная мезенхима, дифференцирующаяся в клетки эпителия.
В, Г. Примитивный клубочковый эпителий сворачивается сначала в структуру, имеющую форму запятой, а затем в S-образную. Д, Е. Проксимальные и дистальные канальцы удлиняются; одновременно формируется структура подоцитов, кровеносные сосуды образуют все больше петель внутри капсулы клубочка, и в конечном счете образуется зрелая сеть клубочковых капилляров. Сосуды клубочков, как полагают, начинают образовываться на ранних стадиях формирования клубочков (В, Г).