Механизмы входящего выпрямления калиевых каналов мембраны клетки
• При потенциал-зависимом блокировании поры происходит входящее выпрямление.
Каким образом различные К+-каналы обеспечивают изменения мембранного потенциала и участвуют в окончании процесса деполяризации мембраны? Большинство ионных каналов обладают способностью к выпрямлению, которое представляет собой изменение ионной проводимости в ответ на изменение заряда мембраны.
Термин входящее (аномальное) выпрямление означает, что ионный канал проводит входящий ток лучше, чем исходящий. В качестве примера приведем семейство К+-каналов входящего выпрямления (Kir), которые эволюционно близки друг к другу. Входящее выпрямление определяется снижением проводимости мембраны для ионов К+ при ее деполяризации и увеличением проводимости при реполяризации ее до более отрицательного мембранного потенциала.
Без входящего выпрямления проводимость канала для К+ была бы постоянной. Однако при недостаточном входящем выпрямлении выходящий поток К+ снижается при более положительном значении мембранного потенциала по сравнению с равновесным потенциалом для К+. При сильно выраженном входящем выпрямлении выходящий поток К+ может полностью блокироваться при более положительном значении мембранного потенциала.
В качестве двух примеров, иллюстрирующих функцию входящего выпрямления, можно привести поддержание отрицательных значений мембранного потенциала покоя и дезактивацию выходящих потоков К+, с целью сохранения внутриклеточной концентрации катиона в течение длительной плато фазы потенциала действия в сердечной мышце. Kir каналы пропускают ионы К+ из покоящейся клетки, но блокируются при деполяризации.
Это имеет физиологический смысл, поскольку деполяризация мембраны обусловлена транспортом в клетку ионов Na+, а одновременный выход положительно заряженных ионов К+ прекратил бы или замедлил деполяризацию мембраны.
Каналы подразделяются в соответствии с их способностью к выпрямлению. Проводимость каналов, обладающих сильной способностью к входящему выпрямлению, резко снижается при достижении значения мембранного потенциала выше определенного уровня. В то же время каналы с недостаточным входящим выпрямлением при этом продолжают транспортировать ионы, хотя и с замедленной скоростью. Каналы входящего выпрямления относятся к классу 2ТМ/1Р или 6ТМ/1Р.
Каналы с сильной способностью к входящему выпрямлению, например Kir, относятся к классу 2ТМ/1Р классу К+-каналов. Различные Kir каналы регулируются различными лигандами, например ионами Mg2+, полиаминами, АТФ или тримерными G белками. Пониженный транспорт К+ через эти каналы при положительном значении потенциала обусловлен блокированием поры со стороны цитоплазмы катионами, например Mg2, или такими лигандами, как полиамины.
В отличие от каналов входящего выпрямления, выходящие каналы, или каналы задержанного выпрямления, относятся к типу 6ТМ/1Р и представляют собой потенциал-зависимые Kv-каналы. Проводимость этих каналов увеличивается по мере деполяризации мембраны при более положительном значении потенциала из-за открытия большего количества каналов. Однако отдельные каналы не обнаруживают способности к выпрямлению. В этом случае говорят об исходящем или задержанном выпрямлении, поскольку активация канала происходит в относительно слабой степени.
Важная роль каналов выпрямления иллюстрируется примером АТФ-чувствительных К+-каналов входящего выпрямления (КАТФ-каналы). Эти каналы являются связующим звеном между обменными процессами и электрической активностью клетки. В b-клетках поджелудочной железы, при увеличении концентрации глюкозы и АТФ, КАТФ-каналы закрываются, и наступает деполяризация мембраны. При этом запускается процесс секреции инсулина. КАТФ-каналы состоят из порообразующих Kir6,2 а-субъединиц и регуляторных субъединиц, функционирующих как метаболический сенсор, контролирующий активность К+-канала.
Мутации в генах, кодирующих белки субъединиц, нарушают функции КАТФ-канала и приводят к постоянной деполяризации мембраны, что сопровождается непрерывной секрецией инсулина, независимо от уровня глюкозы в крови. Такая неконтролируемая секреция инсулина приводит к патологически низкому уровню глюкозы в крови в детском возрасте и к неврологическим расстройствам.
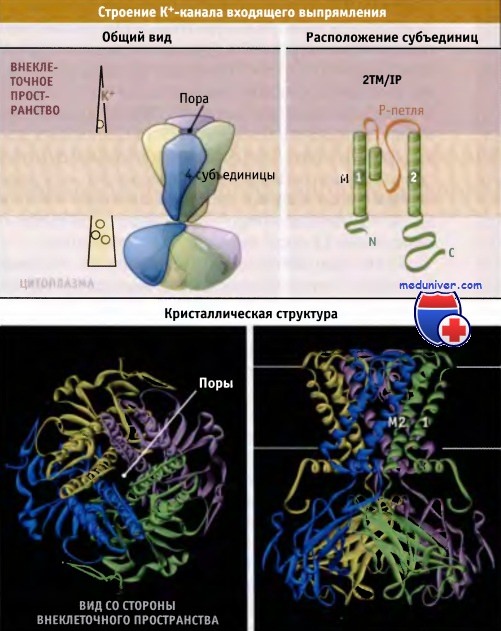
Схематическое строение и кристаллическая структура К+-канала входящего выпрямления (Kir).
Поровый домен по строению близок к домену других К+ каналов.
Кристаллическая структура построена на основании Protein Data Bank file 1P7B.
Вид со стороны внеклеточного пространства по масштабу меньше, чем вид сбоку. Обозначено предполагаемое положение мембраны.
Слева показан градиент К+, существующий в покоящихся клетках животных.