• В клетке белки должны быть локализованы в сигнальных органеллах и мембранах
• Белки непосредственно поступают из цитозоля в несколько типов органелл
• Эндоплазматический ретикулум (ЭПР) представляет собой исходный компартмент, откуда белки вступают на путь секреции. ЭПР приспособлен к выполнению этой функции
• В некоторые органеллы и в плазматическую мембрану белки поступают по секреторному пути
Способность взаимодействовать с окружающей средой и реагировать на ее изменения имеет критическое значение для выживания и нормального функционирования клетки. В окружающей многоклеточный организм среде находятся питательные вещества, факторы роста, гормоны и другие метаболиты, под действием которых клетка может разделиться, вступить на путь дифференцировки или программированной гибели. Клетка должна чувствовать изменения в составе компонентов окружающей среды и реагировать на них соответствующим образом. В зависимости от своих нужд клетки меняют окружение.
Так, они могут секретировать белки, которые используются для построения или разрушения внеклеточного матрикса; могут передавать сигналы соседним клеткам при прямом контакте; а в многоклеточном организме некоторые клетки (например, эндокринные) секретируют гормоны, которые оказывают дистанционный эффект на другие клетки.
Основными посредниками клетки во взаимодействии с окружающей средой являются секреторные и трансмембранные белки. Эти белки синтезируются каждой клеткой, начиная от простейших бактерий до высокодифференцированных и специализированных клеток млекопитающих. Секреторные белки выходят во внеклеточную среду, а трансмембранные белки остаются в плазматической мембране, причем одна их часть контактирует с внешней средой, а другая остается внутри клетки.
Электронная микрофотография содержимого клетки печени.
Показаны характерные для эукариотической клетки органеллы, содержащие мембрану и обладающие различной плотностью.
Видны ядро, митохондрии, лизосомы, пероксисомы и гранулярный эндоплазматический ретикулум.
Фотография любезно предоставлена Дэниелем С. Френдом.
Необходимость полного или частичного транспорта некоторых белков через плазматическую мембрану создает для клетки проблему их отбора. В клетке белки образуются на рибосомах, в цитоплазме. Поэтому должен существовать механизм селективного отбора и транспорта к плазматической мембране только секреторных и трансмембранных белков. У бактерий селективный отбор при транспорте происходит только на белки, входящие в состав плазматической мембраны и не входящие в нее. В клетках эукариот проблема носит более сложный характер.
В то время, как типичная прокариотическая клетка имеет плазматическую мембрану и не обладает внутриклеточными органеллами, клетки эукариот содержат многочисленные структуры, окруженные мембранами, такие как ядра, митохондрии, хлоропласты (у растений), пероксисомы, эндоплазматический ретикулум (ЭПР), аппарат Гольджи и лизосомы.
Для каждой из этих органелл характерен уникальный набор белков. Таким образом, наряду со способностью выделять секреторные и трансмембранные белки, эукариотическая клетка также должна безошибочно узнавать белки органелл. В типичной клетке органеллы занимают примерно половину общего объема. Это означает, что большая часть белков, синтезирующихся в каждый данный момент времени, должна быть селективно локализована в одной из этих органелл.
Каким образом белки локализуются в органеллах? Обычно для направления белка к специфической органелле клетка использует адресные сигналы, которые представляют собой дискретные последовательности аминокислот в составе его первичной структуры. Характер сигнала определяет органеллу, к которой должен быть направлен белок, и эта органелла имеет аппарат специфического распознавания на белке соответствующего сигнала. Если белок синтезировался без адресного сигнала, то он остается в цитозоле.
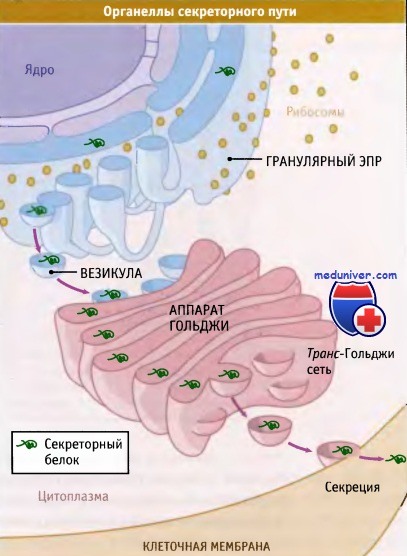
Белки некоторых органелл (митохондрий, хлоропластов, ядра и пероксисом) транспортируются из цитозоля после окончания синтеза на рибосомах. Однако для других органелл (ЭПР, аппарат Гольджи, лизосомы и плазматическая мембрана) процесс протекает более сложно. Процесс транспорта белка через эти органеллы вместе с самими органеллами называется секреторный путь. Непосредственно белки не поступают в аппарат Гольджи, лизосомы и плазматическую мембрану. Вместо этого все белки, подлежащие секреции, или предназначенные для этих содержащих мембраны структур вначале проходят через мембрану ЭПР.
Там они приобретают нативную трехмерную структуру, обычно подвергаются ковалентной модификации, и образуют комплексы с другими белками. Затем эти комплексы попадают в аппарат Гольджи, а оттуда или назад, в ЭПР, или далее, в лизосомы и плазматическую мембрану. Перемещение белков между органеллами секреторного пути происходит с участием мелких везикул, которые отпочковываются от мембраны исходной органеллы и сливаются с мембраной органеллы назначения, высвобождая в нее белковое содержимое.
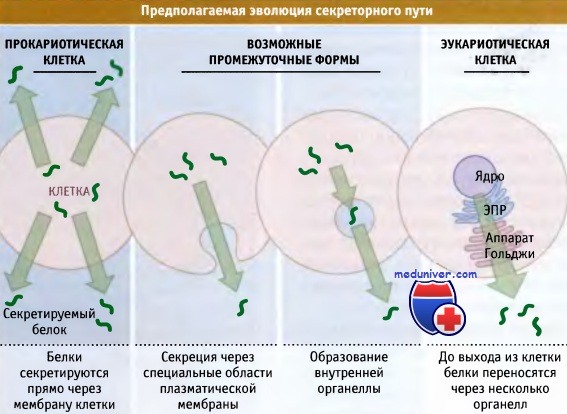
Важнейшая особенность секреторного пути состоит в том, что люмен (внутреннее содержимое) этих органелл по составу напоминает внеклеточное окружение. (Считается, что органеллы возникли в результате инвагинаций плазматической мембраны, специализированной на секреции белка, которые впоследствии превратились в везикулы.) В клетке, секретируемые белки способны принимать нативную структуру, причем это происходит в условиях, близких к тем, в которых они в конце концов должны функционировать.
В отдельных статьях на сайте рассмотрены начальные процессы, которые обеспечивают позиционирование белков на различных органеллах и мембранах внутри клетки. Поскольку почти все белки синтезируются в цитозоле, эти процессы происходят в мембранах органелл, непосредственно импортирующих белки, а именно в мембранах ЭПР, митохондрий, хлоропластов и пероксисом. К числу таких процессов относится узнавание белков, которые, в отличие от других белков цитозоля, должны быть импортированы в органеллу. В дальнейшем эти белки становятся интегральными белками мембраны или транспортируются через нее. Этот процесс известен как транслокация (перенос) белка.
В отдельных статьях на сайте также освещается особая роль, которую играет ЭПР в подготовке белков для экспорта по секреторному пути, и другие функции ЭПР. Вопросы, связанные с транспортом белков по секреторному пути и с их сортировкой в аппарате Гольджи перед адресной доставкой, рассматриваются в отдельных статьях на сайте.
Секреция белков начинается с их адресования и транслокации через мембрану гранулярного эндоплазматического ретикулума.
После формирования нативной структуры и посттрансляционных модификаций белки выходят из ЭПР в везикулах, связывающихся с аппаратом Гольджи.
Большая часть белков переносится из аппарата Гольджи к клеточной поверхности в секреторных везикулах.
Органеллы секреторного пути могли возникнуть при интернализации участка плазматической мембраны,
обладающей функцией секреции белка.
В клетках эукариот белки непосредственно не секретируются через плазматическую мембрану,
а транспортируются в люмен ЭПР, который по составу напоминает внешнее окружение клетки.