• V-АТФазы являются протоновыми насосами, которые состоят из нескольких субъединиц и обладают структурой, близкой к F1F0-АТФ-синтазам
Поскольку функционирование белков зависит от величины pH, этот показатель служит важным параметром, влияющим на протекание многочисленных внутриклеточных процессов. Благодаря метаболизму среда в цитозоле постоянно закисляется и для поддержания постоянного значения pH протоны должны все время удаляться. Поэтому белки, осуществляющие транспорт протонов, необходимы в регуляции pH. Например, pH цитозоля, который играет важную роль в контроле роста клетки и всех матаболических процессов, регулируется такими мембранными переносчиками, как Na+/Н+-обменник. Наряду с этим митохондрии представляют собой некий «резервуар» протонов, образующихся в цитозоле, и F1F0-АТФ-синтаза внутренней мембраны митохондрий использует движущую силу протонов для синтеза АТФ.
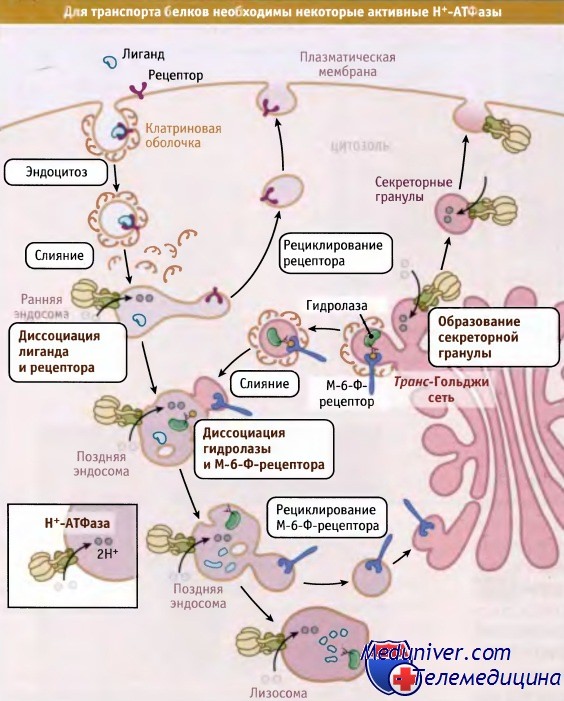
В отличие от цитозоля, в котором поддерживается слабощелочное значение pH, в некоторых клеточных органеллах должна быть кислая среда. К числу таких компартментов относятся органеллы, принимающие участие в процессах эндоцитоза (клатриновые везикулы, эндосомы, лизосомы) и экзоцитоза (секреторные гранулы). В этих органеллах оптимальные значения pH (4,5-6,8) поддерживаются с помощью протоновых насосов вакуолярного типа (V-АТФазы), представляющих собой Н+-АТФазы. Эти АТФазы относятся к группе насосов, которые транспортируют протоны из цитозоля в люмен органелл, используя энергию АТФ. Они играют важную роль в процессах эндоцитоза и внутриклеточного позиционирования.
Для таких процессов, как диссоциация комплексов лиганд-рецептор и возвращение клеточных мембранных рецепторов в плазматическую мембрану, происходящих в ранних эндосомах, необходима кислая среда. Н+-АТФазы также участвуют в подкислении поздних лизосом, которое необходимо для доставки из транс-Гольджи сети лизосомальных ферментов, например содержащих метку манноза-6-фосфата (М-6-Ф). В секреторных гранулах, например в синаптических пузырьках и хромаффинных гранулах, протонный градиент и/или мембранный потенциал, создающийся при действии Н+-АТФаз, обеспечивает движущую силу сопряженного транспорта, а также создание внутри гранул запасы небольших молекул и ионов. Например, захват норадреналина синаптическими пузырьками зависит от протонового градиента, а захват глутамата определяется мембранным потенциалом.
Некоторые вирусы, обладающие внешней оболочкой, например вирусы гриппа, используют кислое значение pH в эндосомальных компартментах для доступа в клетку. Вирусный гемагглютинин при низких значениях pH активируется, что облегчает слияние вируса с эндосомой.
Некоторые V-АТФазы участвуют в везикулярном транспорте.
Закисление среды в эндосомальных компартментах необходимо для диссоциации комплексов белок-рецептор и рециклирования рецепторов.
Наряду с этим, Н+-АТФазы необходимы для образования секреторных гранул.
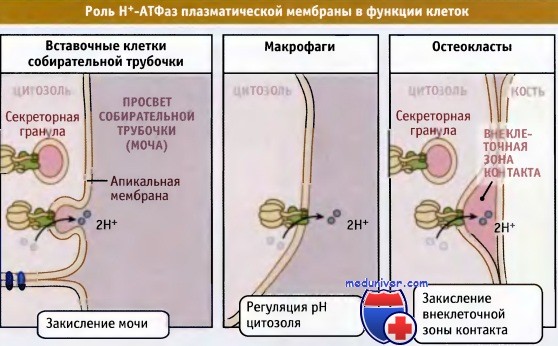
Некоторые Н+-АТФазы находятся в плазматической мембране ряда специализированных клеток, например почек, нейтрофилов и остеокластов. Там они играют важную роль в процессах закисления мочи, поддержания pH цитоплазмы и резорбции костной ткани соответственно. В апикальной мембране эпителиальных вставочных клеток почек находятся в большом количестве Н+-АТФазы, которые активно транспортируют в мочу протоны. Мутации в генах, кодирующих белки этих АТФаз, вызывают нарушения функционирования транспортного механизма, что приводит к пониженной способности почек контролировать pH крови и к развитию почечного и метаболического ацидоза. Нейтрофилы и макрофаги иммунной системы, несмотря на значительное образование кислот в процессе работы, поддерживают нейтральные значения pH цитозоля при участии Н+-АТФаз плазматической мембраны.
В процессе перестройки кости остеокласты присоединяются к костному матриксу, и Н+-АТФазы оказываются в области активной контактной зоны. Закисление этой зоны помогает растворить костный матрикс и активирует гидролазы, участвующие в резорбции кости.
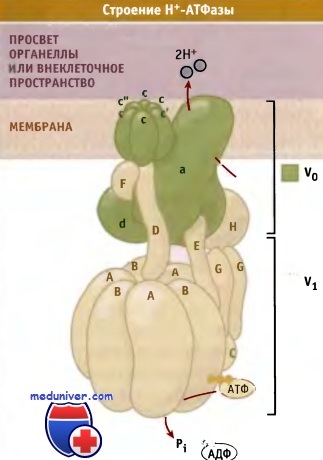
Н+-АТФазы представляют собой белковые комплексы, состоящие из субъединиц и содержащие два функциональных домена V1 и V0. Расположенный в цитозоле домен V1 связывает и гидролизует АТФ. Свободная энергия гидролиза АТФ используется для трансмембранного транспорта протонов, который происходит через мембранный домен V0. Предполагаемая структура фермента построена на основании данных электронной микроскопии, использования метода белковых сшивок и сайт-специфического мутагенеза. Хотя идентифицированы еще не все компоненты насоса, можно предполагать, что цитоплазматический АТФ-связывающий V1-домен с массой 640 кДа состоит из субъединиц А-Н и может быть изображен следующей стехиометрической формулой: A3B3C1D1E1F1G2H1, а домен V0 массой 260 кДа состоит из пяти типов субъединиц: a1d1c"1c'c4.
Н+-АТФазы близки к F1F0-АТФ-синтазам, которые функционируют в обратном направлении и образуют АТФ в митохондриях, хлоропластах и бактериях. Оба типа ферментов имеют два аналогичных домена, F0 или V0, и F1 или V1. Субъединица В домена V1 содержит некаталитический сайт связывания нуклеотида, а субъединица А — каталитический сайт связывания АТФ. Эти сведения были получены на основании экспериментов по сайт-зависимому мутагенезу и по использованию блокаторов сульфгидрильных групп. Поскольку оба фермента близки друг к другу по строению, механизм функционирования Н+-АТФаз, вероятно, аналогичен хорошо изученному механизму действия F-АТФ-синтаз.
В специализированных клетках Н+-АТФазы плазматических мембран обладают различными функциями.
По-видимому, F-АТФ-синтазы функционируют как роторные двигатели, в которых протонный градиент приводит во вращение кольцо с-субъединиц. Вращение происходит относительно субъединицы а домена F0. Энергия вращения через центральный выступ γ и субъединицу ε передается на кольцевую структуру и используется субъединицами а и b для синтеза АТФ и высвобождения. Субъединицы с содержат остатки аспарагиновой кислоты, которые играют важную роль в транспорте протонов, а субъединица а обеспечивает протонам доступ к кольцу с-субъединиц.
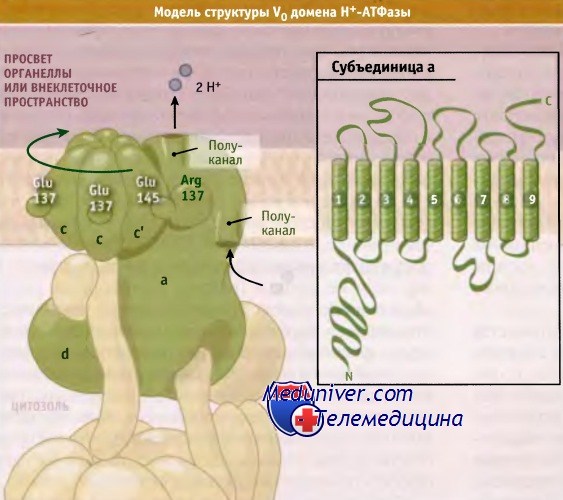
Аналогичная модель, описывающая функционирование Н+-АТФаз, представлена на рисунке ниже. Согласно этой модели, энергия, которая высвобождается при гидролизе АТФ доменом V1, обеспечивает вращение с-кольца и перенос протонов. Субъединица а, входящая в состав домена V0, переносит протоны через девять предполагаемых трансмембранных сегментов, расположенных в ее С-концевой области. Положительно заряженный остаток аргинина в субъединице а стабилизирует отрицательно заряженный остаток глутамата в одной из субъединиц с до момента ее протонизации.
В субъединице а для транспорта протона и обратимого протонирования остатка глутамата, который высвобождает его при взаимодействии с остатком аргинина субъединицы с, используются две полуканальные структуры. Электростатическое притяжение аргининового остатка к непротонированному глутамату следующей с-субъединицы позволяет с-кольцу вращаться, что приводит каждую субъединицу с в контакт с субъединицей а. Этот механизм обеспечивает односторонний транспорт протонов за счет энергии гидролиза АТФ.
Активность H+-АТФаз регулируется по нескольким механизмам. Один механизм включает регулируемое позиционирование и слияние внутриклеточных везикул, содержащих протонные насосы, с плазматической мембраной. Например, во вставочных клетках почек секреторные везикулы, содержащие Н+-АТФазы, в ответ на снижение внутриклеточного pH, обратимо сливаются с апикальной мембраной. Согласно второму механизму, образование дисульфидных связей между консервативными остатками цистеина, расположенными неподалеку от каталитического центра субъединицы а, приводит к обратимому ингибированию активности АТФазы. Третий механизм постулирует, что в определенных условиях обратимая диссоциация комплекса на домены V1 и V0 служит важным механизмом регуляции активности Н+-АТФазы.
Модель, описывающая строение Н+-АТФаз.
Эти ферменты транспортируют протоны из цитозоля в просвет везикул или во внеклеточное пространство посредством интегрированного в мембрану Vо домена.
Цитоплазматический V1 домен расходует энергию гидролиза АТФ на транспорт протонов.
Модель, описывающая вращение с-кольца и движение протонов через V0-домен
(гидролиз АТФ в V1-домене, обеспечивающий это движение, не показан).
В рамке показана схема предположительного строения субъединицы а домена V0,
построенная на основании данных по мутагенезу и при использовании химических модификаторов.