• Деление клеток не является непрерывным процессом и контролируется внешними стимулами и наличием питательных веществ
• Клетки узнают химические сигналы в своем окружении
• Внешние сигналы могут вызывать в клетке биохимический ответ, который приводит или к ее вхождению в цикл, или к остановке цикла в фазе G1/G0
Здоровые клетки пролиферируют только при наличии подходящих условий. В случае одноклеточных организмов, необходимые условия для пролиферации создаются при наличии питательных компонентов. Для клеток многоклеточных необходимость вступления в цикл деления определяется дополнительными факторами клеточного окружения. Такие внешние сигналы могут обладать активирующими или ингибирующими свойствами. Каким образом все эти сигналы реализуются и интегрируются в машину клеточного цикла?
Пептиды и гормоны, которые стимулируют пролиферацию, называются ростовыми факторами. Эти факторы находятся в сыворотке крови, что и объясняет ее использование в средах для культивирвоания клеток in vitro. Одним из первых идентифицированных ростовых факторов сыворотки был пептид, названный тромбоцитарным фактором роста (PDGF). Этот фактор выделяется тромбоцитами при свертывании крови и способствует быстрой пролиферации клеток, необходимой для заживления ран.
PDGF и другие ростовые факторы связываются со специфическими рецепторами на поверхности клеток-мишеней. Эти рецепторы связаны с процессами внутриклеточной системы передачи сигнала, которые индуцируют пролиферацию.
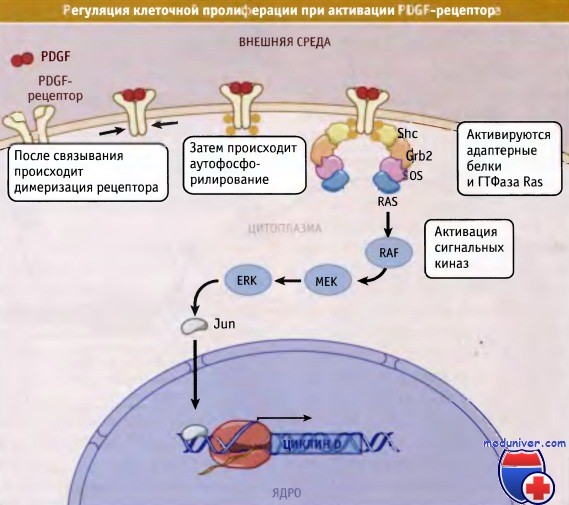
Каким образом ростовые факторы вызывают стимуляцию активности CDK, необходимую для коммитирования вступления клетки в S-фазу и митоз? После связывания фактора роста и активации рецепторов происходит множество биохимических процессов, и в клетке образуются вторичные мессенджеры. В конце концов, как показано на рисунке ниже, это приводит к изменениям экспрессии генов. Набор генов, которые экспрессируются вскоре после добавления к клеткам сыворотки, называют генами раннего ответа.
Многие из них, кодирующие факторы транскрипции, например Fos, Jun и Мус, которые активируют другие гены, называются замедленными генами раннего ответа. Одним из таких генов является ген, кодирующий G1-циклин, циклин D. Хотя активация CDK в ответ на воздействие факторов роста также требует прохождения других процессов, стимуляция экспрессии циклина D представляет собой важнейший механизм продвижения клеток через переход G1/S.
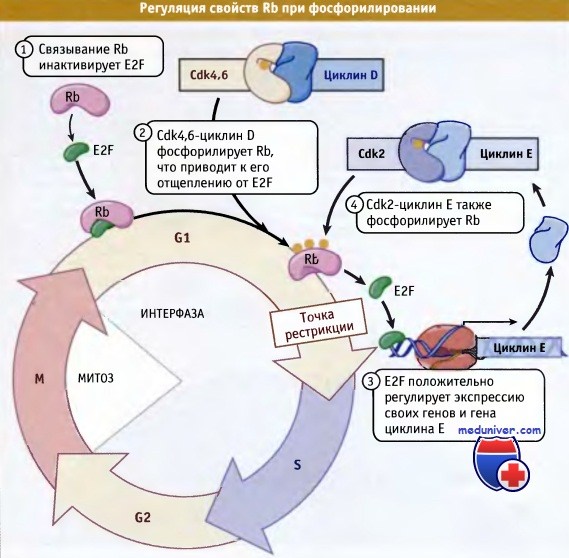
CDK и связанные с ними G1-циклины способствуют прохождению клетки через точку рестрикции за счет фосфорилирования супрессора опухоли ретинобластомы (Rb), который связывается с фактором транскрипции E2F и ингибирует его. Фосфорилирование Rb комплексом G1 CDK-циклин (главным образом, циклин D, связанный или с Cdk4 или с Cdk6) приводит к его отщеплению от E2F. Активный E2F теперь стимулирует свою собственную экспрессию, а также экспрессию других генов. К этим генам относятся циклин Е, белки инициации репликации, и другие белки, участвующие на поздних стадиях клеточного цикла.
Cdk2-циклин Е также фосфорилирует Rb, что усиливает эффект первоначального выхода Rb из комплекса с E2F. Таким образом, небольшая исходная активность CDK может привести к существенному увеличению уровня и активности G1 CDK-циклиновых комплексов. Процессы регуляции Rb и E2F CDK-циклиновым комплексом схематически представлены на рисунке ниже. Утрата функциональной активности Rb играет важную роль в развитии опухолей.
Таким образом, общие характерные свойства циклин-зависимых киназ проявляются на разных стадиях клеточного цикла. Однако, как мы увидим, при репликации эта зависимость клеточного цикла связана с дерегуляцией отдельных точек начала репликации ДНК на хромосомах.
Связывание лиганда с рецептором на наружной мембране клетки приводит к димеризации и к активации рецептора.
В представленном примере рецептор Tyr-киназы, рецептор PDGF, подвергается внутримолекулярному фосфорилированию,
что активирует адаптерные белки Shc, Grb2 и SOS. Эти белки активируют ГТФазу, называемую Ras.
В результате происходит активация сигнального каскада киназ, включающего киназы RAF, MEK и ERK,
которые, в конце концов, активируют транскрипцию.
При этом индуцируется экспрессия генов, регулирующих пролиферацию и прохождение клеток через точку рестрикции, что обеспечивает вступление клетки в цикл деления.
Связывание Rb фактором транскрипции E2F приводит к инактивации последнего.
Когда Rb фосфорилируется, он не может связываться с фактором и таким образом обеспечивает ему возможность положительно регулировать экспрессию многих генов,
включая свой собственный, а также ген циклина Е.
Затем Cdk2-циклин Е может фосфорилировать большее количество Rb, что приводит к петле усиления.
Когда накапливается достаточно Cdk-циклина Е, клетка проходит точку рестрикции.