• Незавершенный и/или дефектный процесс репликации ДНК активирует точку контроля клеточного цикла
• Дефекты ДНК активируют другую точку контроля, которая содержит компоненты, общие с точкой контроля за репликацией
• Точка контроля за повреждениями в ДНК блокирует цикл в определенных стадиях, в зависимости от стадии цикла, в которую произошло повреждение
В предыдущей статье на сайте мы дали общие представления о точках контроля. Сейчас мы рассмотрим конкретные точки, которые защищают клетку. Дефекты ДНК представляют для клетки большую угрозу, и состояние генома постоянно контролируется. Существует также специальная точка контроля, которая функционирует в S-фазе и контролирует процесс репликации ДНК.
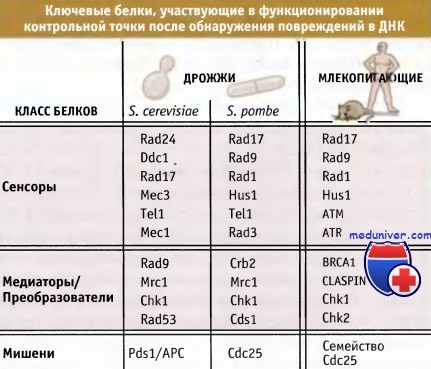
Сейчас известно, что в реакции клетки на воздействие факторов, повреждающих ДНК или тормозящих ее репликацию, участвует множество белков, и некоторые из них перечислены на рисунке ниже.
Молекулярные основы узнавания дефектов являются наименее понятным аспектом функционирования точек контроля за повреждениями и репликацией ДНК. Тем не менее очевидно, что изменения структуры ДНК, такие как образование локальных одноцепочечных участков (ssДHK) или двунитевых разрывов (ДР), узнаются точкой контроля за повреждениями.
Образование участков с неправильной структурой приводит к мобилизации и активации близких по свойствам протеинкиназ, обладающих консервативной структурой, ATR (и ее адаптера ATRIP) и АТМ.
У дрожжей и высших эукариот области ssДHK вызывают ATR-зависимый ответ, в то время как ДР приводят к процессам, зависимым от АТМ. Исследования на почкующихся дрожжах показали, что присутствие одного ДР, и последующее образование участков ssДHK или образование ssДHK у мутантов cdc13, контролируются точкой, зависимой от Mec1 киназы, которая является гомологом ATR.
Было показано, что на каждом этапе функционирования контрольной точки
некоторые эволюционно-консервативные белки выполняют определенные функции.
С другой стороны, у-облучение клеток млекопитающих, при котором образуются ДР, вызывает всплеск активности АТМ киназы и последующее увеличение активности ATR, вероятно, связанное с образованием участков ssДHK в местах образования ДР. Несомненно, что АТМ и ATR киназы играют ключевую роль в узнавании дефектов в ДНК и их ликвидации, и процессы, в которых участвуют обе киназы, в значительной степени перекрываются.
Не исключено, что даже при задержке продвижения репликативной вилки точка контроля за репликацией узнает ssДHK в этой области, что приводит к активации ATM/ATR киназ.
Каким образом узнаются дефекты хромосом? Исследования с использованием электронной микроскопии показали, что АТМ киназа может непосредственно связываться с ДНК на концах ДР. Вероятно, ATR киназа преимущественно связывается с УФ-индуцированными дефектами ДНК. Затем связывание приводит к увеличению активности киназы и к дальнейшему распространению сигнала.
Как происходит активация АТМ? Эта киназа в неактивносм состоянии существует в виде гомодимера. При повреждении ДНК запускается процесс аутофосфорилирования фермента, который сопровождается диссоциацией молекулы и высвобождением активных мономеров, которые фосфорилируют большое количество белков-мишеней.
Наряду с ATM/ATR киназами, в узнавании повреждений ДНК независимо участвуют эволюционно-консервативные белки Rad17-RFC и комплексы 9-1-1. Семейство белков Rad17 взаимодействует с компонентами фактора репликации С и образует комплекс точки контроля, обладающий способностью связываться с ДНК. Затем комплекс Radl7-RFC связывает комплекс 9-1-1 (который у человека состоит из трех белков, называемых Rad9, Rad1 и Hus1) с дефектами ДНК. Комплекс 9-1-1 образует кольцевую структуру, напоминающую «скрепку» PCNA, участвующую в репликации ДНК.
Несмотря на то что все эти белки независимо узнают дефекты ДНК и связываются по этим местам, все они необходимы для функционирования точек контроля за повреждениями ДНК. Вопрос, каким образом в местах дефектов ДНК осуществляется взаимодействие между ними, еще является предметом интенсивных исследований.
Медиаторы точек контроля представляют собой следующую группу белков, необходимых для узнавания дефектов в ДНК, передачи сигнала и репарации ДНК. Эти белки действуют после того, как дефект был обнаружен, и представлены на рисунке ниже. Для этих белков нельзя провести четкой границы между их функционированием в процессах точки контроля и репарации ДНК, поскольку некоторые из них участвуют в обоих процессах. Также медиаторы могут обеспечивать поддержку систем узнавания дефектов ДНК за счет стабилизации мультибелковых сигнальных комплексов.
Основными передатчиками точек контроля за повреждениями и репликацией ДНК являются Chk1 и Chk2 киназы. Эти ферменты непосредственно фосфорилируют компоненты машины клеточного цикла и блокируют их активность.
Идентичные дефекты структуры ДНК могут задержать или остановить клетку в той или иной фазе цикла, в зависимости от стадии, в которой они были узнаны точкой контроля. Например, если повреждения в ДНК обнаружены в стадии G1, то прохождение клетки по циклу задерживается в точке перехода G1/S. Если тот же дефект обнаруживается в течение G2, блокируется вхождение клетки в митоз.
Существует три самостоятельных точки контроля, которые блокируют продвижение клетки по циклу при появлении дефектов в ДНК или задержке репликации: точка контроля G1/S, точка контроля в S-фазе и точка на границе G2/M. Однако для этих точек характерно присутствие одних и тех же компонентов.
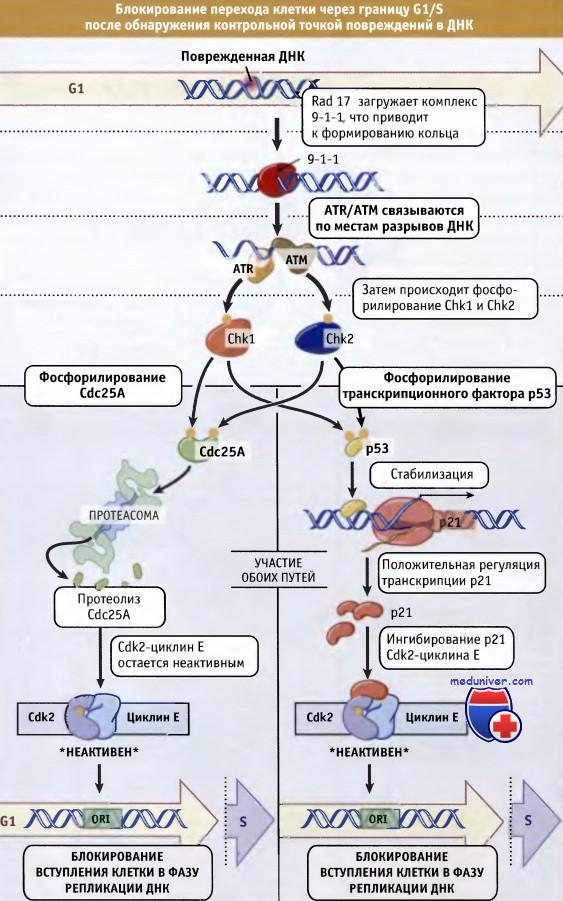
Точка контроля G1 в клетках млекопитающих исследована достаточно подробно. Она задерживает вступление клетки в S фазу, ингибируя активность G1 CDK-циклинового комплекса. Ингибирование может происходить даже после прохождения клеткой точки рестрикции. Каким образом это достигается? Блок G1 достигается по меньшей мере двумя путями. Один путь обеспечивается ингибированием активации G1 CDK-циклиновых комплексов.
Активация ATM/ATR киназ приводит к фосфорилированию и активации эффекторных киназ Chk2 и/или Chk1. Затем эти киназы фосфорилируют Cdc25A, что приводит или к выходу ее из ядра, или к деградации по убиквитин-зависимому механизму. В результате, комплекс Cdk2-циклин Е поддерживается в неактивном состоянии за счет фосфорилирования тирозинового остатка.
Другой путь, приводящий к блоку перехода G1/S при появлении дефектов ДНК, включает активацию транскрипции CKI. После активации точки контроля происходит активация ATM/ATR и фосфорилирование р53-Этот белок представляет собой фактор транскрипции и супрессор опухолевого роста. Фосфорилирование р53, которое происходит под действием ATM/ATR, предотвращает его деградацию, и белок накапливается в ядре клетки. Это вызывает активацию транскрипции генов, находящихся под контролем р53, таких как р21.
Последний представляет собой CKI и ингибирует CDK-циклиновые комплексы. Таким образом, ингибирование CDK-циклиновых комплексов, необходимое для репликации ДНК, происходит двумя разными путями. На рисунке ниже схематически представлены процессы функционирования G1 точки контроля, которые запускаются в ответ на возникновение дефектов в ДНК.
Характер ответа, который генерируется точками контроля за повреждениями ДНК в S-фазе и на границе G2-M, принципиально близок к таковому для точки контроля G1. Точка контроля в S-фазе в основном обеспечивает задержку активации поздних точек начала репикации, когда узнаны дефекты ДНК, с тем чтобы предотвратить разборку репликативной вилки. Так же как и в случае точки контроля в G1-фазе, ключевыми участниками являются ATM/ATR киназы и эффекторные киназы Chkl и Chk2, которые обеспечивают ингибирование Cdc25A и CDK-циклиновых комплексов. Однако, наряду с этим, активные АТМ/ATR киназы фосфорилируют белки репликации, что вызывает задержку ее наступления.
Например, ATR-ATRIP ингибирует киназу Cdc7-Dbf4 и тем самым предотвращает связывание белка Cdc45, необходимого для репликации. За счет этого подавляется инициация процесса в S-фазе. Еще одной важной мишенью киназ точки контроля является экзонуклеазный комплекс, который действует на двунитевые разрывы ДНК и называется MRN (Mre11/Rad50/Nbs1).
Если точка контроля в S-фазе не функционирует, то синтез ДНК продолжается при наличии в ней дефектов. Этот процесс обозначается как радиоустойчивый синтез ДНК (RDS). Интенсивность радиоустойчивого синтеза ДНК увеличивается, если контрольные процессы клеточного цикла (ATM/ATR-Chkl/Chk2-Cdc25A) и остановки репликации не функционируют. Это позволяет предполагать, что события, которые инициируются дефектами в ДНК в S-фазе, происходят параллельно. Процессы характерные для точки контроля в S-фазе представлены на рисунке ниже.
Точка контроля G2-M аналогична точкам в S- и Gl-фазе в том, что активация цепочки процессов с участием ATM/ATR-Chk1/Chk2 приводит к ингибированию комплекса CDK-циклин, в данном случае комплекса CDK1-циклин В. Опять-таки, в этом случае мишенями для Chk1 и Chk2 киназ служит белок семейства Cdc25. Для точки контроля G2-M характерно, что активность Cdc25C и Cdc25A подавляется за счет их ингибирования и/или деградации.
Методы прижизненного исследования клеток и использование зеленого флуоресцирующего белка обеспечили дополнительную возможность выяснить, когда и в течение какого времени действует каждый компонент процессов функционирования контрольной точки. В местах повреждений ДНК или задержки репликативных вилок накапливается много белков, необходимых для функционирования контрольных точек.
Предполагая, что эти места соответствуют сигнальным сайтам для контрольных точек, исследователи пытаются установить порядок появления на них соответствующих белков, продолжительность их нахождения там, и выяснить зависимость этих параметров от присутствия других белков. Возможность визуализации мест накопления белков также представляет собой удобный экспериментальный подход, позволяющий проверить наличие повреждений в ДНК и оценить степень их тяжести.
Фосфорилирование С-терминального хвоста гистона Н2АХ под действием ATM/ATR с образованием у-Н2АХ может являться сигналом наличия дефекта в ДНК. Хотя мобилизация некоторых белков точек контроля не зависит от у-Н2АХ, постоянное их присутствие в местах повреждений и развитие последующих событий, вызванных точкой контроля, от него зависит. Поэтому у-Н2АХ может представлять собой сигнальный центр для контрольной точки и репаративных белков, обеспечивающий функционирование процесса репарации ДНК и контроль за ним.
Точки контроля за повреждениями ДНК играют критическую роль в поддержании стабильности генома. Нарушение функций белков, участвующих в процессах контроля, может приводить к накоплению мутаций и к развитию рака. Такое генетическое заболевание человека, как атаксия-телеангиэктазия (А-Т), вызывает разрушение мозжечка, развитие иммунодефицита, повышение радиочувствительности и нестабильности генома, а также предрасположенность к развитию злокачественных опухолей. Клонирование мутантного гена от больных А-Т показало, что он представляет собой АТМ. Была установлена важная роль этого белка в обеспечении стабильности генома.
Частичная утрата белком ATR своих функций приводит к редкому аутосомному рецессивному заболеванию — синдрому Секеля.
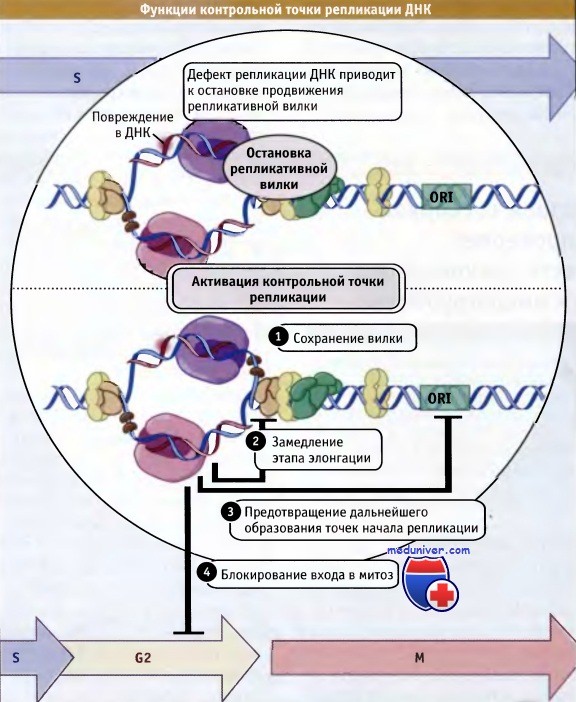
Как функционирует точка контроля за репликацией ДНК и отличается ли она от точек контроля за повреждениями, образующимися в этой молекуле? Исследования на многоклеточных показали, что активация точки контроля за репликацией происходит в ответ на остановку продвижения репликативных вилок. Эта точка не активируется, если ДНК реплицировалась неполностью. Снижение пула dNTP под действием таких веществ, как оксимочевина, или мутации по генам, кодирующим синтез ферментов репликации, приводит к ее остановке и к активации точки контроля. Однако такая активация не происходит у клеток, не способных к сборке репликативной вилки.
После запуска процессов точки контроля за репликацией могут произойти следующие четыре события:
• предотвращение активации дальнейших точек начала репликации;
• замедление этапа элонгации репликации;
• поддержание состояния блокировки репликативных вилок, и
• ингибирование входа клетки в митоз.
Эти события схематически представлены на рисунке ниже. Многие компоненты участвуют в процессах контроля за репликацией и за дефектами в ДНК. Например, главные контрольные киназы — ATR и ее гомологи и Chkl с гомологами, сигнализируют белкам из группы Cdc25 о необходимости ингибирования активности комплекса CDK-циклин В и входа клетки в митоз. Фактически эти киназы участвуют в трех из перечисленных процессов. Однако они не требуются для замедления этапа элонгации при репликации ДНК, и этот аспект функционирования точки контроля исследован еще недостаточно.
После обнаружения повреждений в ДНК в G1-периоде, в клетке активируется контрольная точка с участием процессов, зависимых от ATM и ATR.
Это может осуществляться, по меньшей мере, двумя путями. Один путь предполагает фосфорилирование Cdc25A фосфатазы.
Это вызывает ее протеолиз, и Cdk2-циклин Е поддерживается в неактивном состоянии, поскольку на остатке Tyr остается фосфатная группа.
Согласно второму пути, происходит фосфорилирование транскрипционного фактора р53, который тем самым стабилизируется.
Стабилизация р53 приводит к положительной регуляции транскрипции белка р21.
В свою очередь, это приводит к накоплению р21 CKI, который затем связывается с Cdk2-циклин Е и блокирует его.
В блокировании перехода клетки через границу G1/S участвуют оба пути.
При незавершении или при ошибке репликации ДНК контрольная точка S-фазы генерирует блокирующий сигнал.
Этот сигнал предотвращает вхождение клетки в митоз до момента коррекции ошибки.
Дефект репликации ДНК вызывает остановку репликативных вилок.
Присутствие этих структур активирует контрольную точку репликации,
которая функционирует для того, чтобы (3) предотвратить дальнейшее образование точек начала репликации, (2) замедлить этап элонгации,
(1) сохранить вилку и (4) заблокировать вход клетки в митоз.