• Репликация происходит после прохождения клеткой точки рестрикции или START
• Репликация регулируется поэтапно и скоординирована с наступлением митоза
• Репликация происходит в точках инициации, которые могут обладать особой первичной структурой, специфическим положением, или располагаться в ДНК на определенных расстояниях
• Инициация происходит только в разрешенных точках, способных к репликации
• Осуществив свои функции, до наступления следующего цикла, точки начала репликации не могут использоваться повторно
Когда клетки проанализировали состояние окружающей среды и приняли решение вступить в цикл деления, они проходят точку G1/S и начинают репликацию ДНК. Как клетки объединяют и активируют факторы, необходимые для репликации ДНК? Какие механизмы контроля гарантируют, что клетка лишь однажды реплицировала ДНК, и только один раз в цикле?
Хотя пока невозможно дать полные ответы на эти вопросы, в результате идентификации и анализа последовательности дрожжевых хромосомных ДНК, способных к независимой репликации, было получено много информации о самом процессе репликации ДНК Эти последовательности в хромосоме, называемые автономно реплицирующиеся последовательности (ARS), являются частью точек начала репликации. Точка инициации или начала (ориджин репликации) представляет собой участок последовательности ДНК, на котором начинается репликация.
У почкующихся дрожжей, но не у большинства других организмов, точки начала репликации представлены небольшими консенсусными последовательностями. У сливающихся дрожжей эти точки занимают большие участки ДНК, богатые АД парами, но не отличаются какой-то особой структурой. У остальных эукариот точки начала репликации расположены случайно, в соответствии с распределением по геному белков, неспецифически связанных с ДНК.
Для того чтобы гарантировать своевременную дупликацию генома, на хромосоме должно быть достаточное количество точек начала репликации. У бактерий для репликации единственной кольцевой хромосомы необходима лишь одна точка начала, однако у эукариот, имеющих большой геном, распределенный по многим линейным хромосомам, должно быть много точек начала репликации. У почкующихся дрожжей, величина генома которых составляет около 13 Мб, в 16 хромосомах находится примерно 400 точек начала репликации.
Это создает несколько проблем, связанных с регуляцией процесса репликации. Функционирование точек начала репликации должно быть скоординировано с клеточным циклом таким образом, чтобы репликация начиналась только в течение S фазы. Должна быть полная уверенность в том, что репликация завершилась до перехода клетки в митоз. Каждая из точек начала репликации должна функционировать только один раз, с тем чтобы гарантировать, что ДНК реплицируется лишь один раз за цикл.
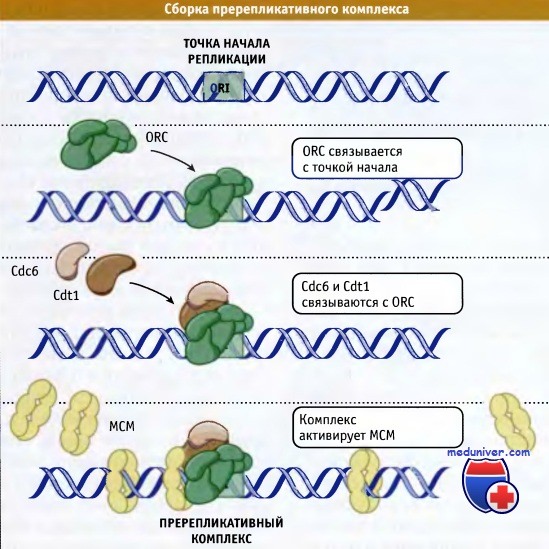
В продолжение всего клеточного цикла 0RC связан с точкой начала репликации на хромосоме.
В течение короткого промежутка времени, от поздего митоза до G1, с точкой начала также связываются белки, активирующие репликацию,
Cdc6 и Cdt1, что, в свою очередь, активирует гексамерный МСМ геликазный комплекс (МСМ2-7).
Этот этап завершает снятие блока репликации и сборку пре-RC.
Точки начала репликации связывают факторы, необходимые для активации репликации и инициации синтеза ДНК. Инициация репликации ДНК происходит только в тех точках, которые содержат связанные факторы и которые поэтому относятся к разрешенным точкам. Однако в каждом раунде репликации ДНК в процессе участвует ограниченный набор потенциальных начальных точек, присутствующих в хромосомах. Более того, по мере активации в разное время различных разрешенных начальных точек, события инициации также происходят через различные интервалы времени. Например, некоторые точки активируются в ранней S-фазе, а другие переходят в это состояние позже.
Неизвестно, чем задается этот временной фактор, но похоже, что расположение специфической точки начала репликации на хромосоме определяет время ее наступления.
Ддя того чтобы репликация началась, в точке начала должен сформироваться пререпликативный комплекс (pre-RC). В результате проведения генетических исследований на дрожжах и биохимических экспрериментов на экстрактах яйцеклеток Xenopus, выяснилась картина сборки pre-RC. Процесс начинается со связывания с ДНК комплекса из шести белков, который носит название комплекса, распознающего область начала репликации (origin recognition complex, ORC). Этот комплекс помечает потенциальную точку начала репликации, однако его недостаточно для активации. Он служит платформой для связывания еще двух консервативных белков: Cdc6, который относится к семейству ААА+АТФазы, и Cdt1. (Многие белки, обладающие АТФазным доменом, для выполнения работы используют энергию АТФ.)
Затем к ним присоединяется комплекс поддерживающий минихромосому (minichromosome maintenance complex, МСМ), представляющий собой кольцевую структуру, состоящую из шести родственных белков, которые также относятся к большому семейству ААА+АТФаз. Комплексы МСМ присутствуют в избытке и распространяются за пределы точки начала репликации. После присоединения МСМ, ORC и Cdc6 становятся необязательными компонентами и pre-RC переходит в состояние способное к активации. Порядок событий, происходящих при сборке pre-RC в точках начала репликации, схематически представлен на рисунке ниже.
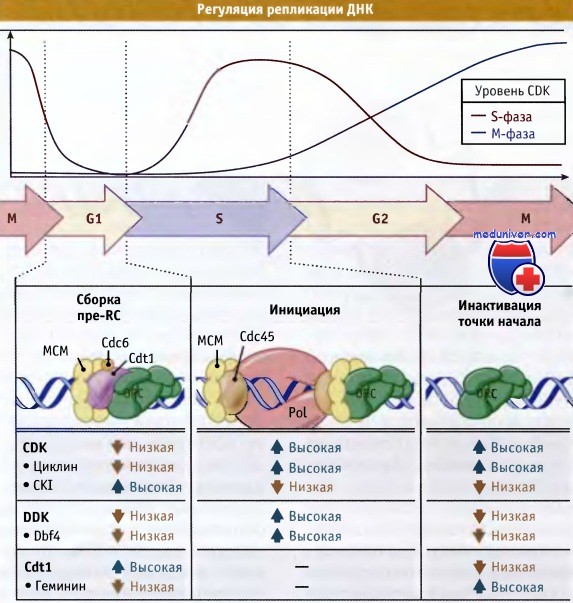
Сборка pre-RC ограничена промежутком между концом М-фазы и ранней S-фазой, что объясняется следующими причинами. Во-первых, в клетке уровень белка Cdc6 контролируется таким образом, что он присутствует только в этот промежуток времени. В отсутствие белка Cdc6, МСМ белок не связывается с точкой начала репликации. Во-вторых, у Метазоа, белок Cdt1 негативно регулируется другим белком, геминином, который блокирует его активность во всех периодах, за исключением окна в G1. Наконец, сборка самого pre-RC ограничивается активностью митотического CDK-циклинового комплекса.
Субстратами для этого комплекса являются субъединицы ORC, Cdc6 и МСМ. При фосфорилировании Cdc6 инактивируется, а фосфорилирование белка МСМ в S-фазе вызывает его отщепление от ДНК. Поэтому pre-RC может сформироваться только при низкой активности CDK-циклинового комплекса. Это характерно для промежутка между уровнями высокой активности митотического комплекса CDK-циклина в М-фазе, и в S-фазе, когда она снова увеличивается, способствуя активации точки начала репликации.
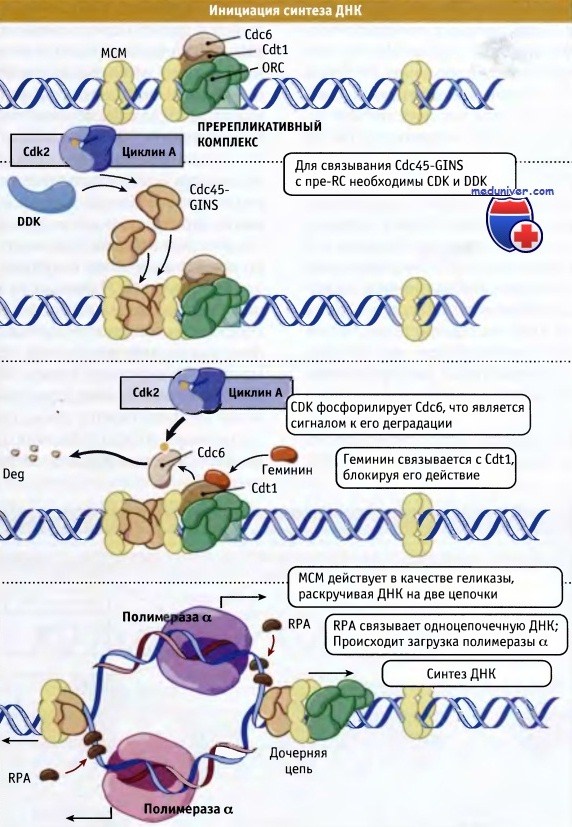
Каким образом точка начала репликации переходит из пререпликативного в репликативное состояние? Для такого перехода необходимо формирование многих дополнительных белковых комплексов, и процесс находится под контролем двух киназ, комплекса CDK-циклин и Cdc7-Dbf4 (DDK). Таким образом, активность CDK-циклинового комплекса координирует процессы репликации и клеточного цикла. При этом координация может носить как негативный (предотвращая сборку pre-RC и, таким образом, повторное функционирование точек начала репликации), так и позитивный характер (промотируя активацию точек начала репликации). К числу вопросов, ожидающих своего ответа, относится вопрос относительно субстратов киназ, промотирующих инициацию репликации.
Если комплексы CDK-циклин обеспечивают координацию процессов во всем цикле, то DDK действует на уровне отдельных точек, инициирующих синтез ДНК. К числу наиболее известных субстратов этой киназы относятся сами МСМ-белки. Интересно, что точечная мутация в гене Mcm5 отменяет необходимость присутствия DDK. Это позволяет предполагать, что фосфорилирование приводит к изменению струтуры белка МСМ, что служит причиной инициации репликации. Впрочем, имеющиеся данные свидетельствуют о том, что эти изменения крайне незначительны. На показаны контрольные процессы репликации ДНК с участием CDK и DDK.
Вероятно, лимитирующим процессом инициации репликации в отдельных начальных точках является связывание белка Cdc45, для которого необходимы как CDK-циклиновый комплекс, так и DDK. Связывание этого белка, которое сопровождается образованием дополнительного комплекса, называемого GINS, приводит к раскручиванию спирали ДНК в начальной точке, за счет активации комплекса МСМ, действующего как хеликаза. Таким образом, МСМ превращается из фактора сборки, участвующего в формировании pre-RC, в фермент хеликазу, который является частью комплекса элонгации. Раскручивание двойной спирали ДНК в точке начала репликации приводит к образованию однонитевой ДНК, которая связывает специфический белок RPA, относящийся к группе белков, связывающихся с однонитевой ДНК (ssDNA binding proteins).
В свою очередь, белок RPA способствует связыванию комплекса праймаза/ДНК-полимераза альфа, который инициирует синтез ДНК. Комплекс МСМ и Cdc45 движется вдоль ДНК, формируя расширяющуюся репликативную вилку, которая образует большую реплисому, включающую в основном ДНК-полимеразу 6, а не полимеразу а. Процессы инициации репликации ДНК представлены на рисунке ниже. В поддержании функционирования реплисомы и защите ДНК в области репликативной вилки от повреждений участвуют контрольные точки и системы репарации ДНК.
Как только МСМ отошли от точки начала репликации, она считается «использованной» и не может активироваться повторно до окончания следующей М-фазы, пока в начальной точке снова не сформируется pre-RC. По мере прохождения S-фазы, МСМ удаляются из хроматина. Наряду с этим, при движении репликативной вилки устанавливаются связи, соединяющие вместе до наступления митоза вновь образованные сестринские хроматиды. Таким образом, завершение S-фазы связано с формированием структур, необходимых для правильной сегрегации хромосом в митозе, что свидетельствует об образовании связей между различными фазами клеточного цикла.
Функция точек начала репликации также регулируется на уровне целой хромосомы. Целесообразно напомнить, что в клетке ДНК с помощью нуклеосом упакована в хроматин, который накладывает на нее ряд структурных ограничений. Это обстоятельство может влиять на временную организацию репликации; например не на всех точках начала в S-фазе репликация происходит в одно и то же время. В некоторых случаях относительное время наступления активации точек начала репликации может определяться не самой точкой, а ее расположением на хромосоме. Точки, расположенные поблизости от транскрипционно-активного эухроматина, инициируются раньше, чем расположенные вблизи транскрипционно-неактивного гетерохроматина, которые обычно инициируются в поздней S-фазе.
Например, точки, расположенные вблизи теломерных областей хромосом (которые обычно транскрипционно неактивны), реплицируются в конце S-фазы. Это общее правило подтверждается изящными экспериментами, выполненными на почкующихся дрожжах. В этих экспериментах перемещение поздней точки начала репликации в эухроматиновую область приводило к ее ранней репликации, и наоборот Однако время наступления начала репликации контролируется также внутренними факторами, присущими самой точке, и роль структуры хроматина и ядра в динамике репликации выяснена недостаточно.
Таким образом, процесс дупликации генома требует интеграции разнообразных сигналов, связывающих вместе регуляторные системы всего клеточного цикла, контролирующие состояние клетки (CDK), и специфические белки хроматина, которые регулируют индивидуальные cis-сайты. Скоординированное действие CDK и DDK демонстрирует, как различные типы киназ обеспечивают выработку конвергентного сигнала, управляющего прохождением клетки по циклу.
Сборка пре-RC происходит в поздней М-фазе и в G1, когда активность CDK и DDK находится на низком уровне.
По мере роста их активности начинается инициация синтеза ДНК.
После инициации npe-RC разбирается и может быть собран повторно, только когда в конце митоза активность CDK снова понизится.