С открытием нуклеосом, оказавшим революционизирующее влияние на состояние наших знаний о строении хроматина, оформились два одновременных направления дальнейших исследований:
• Структурный анализ, описывающий строение самих нуклеосом и способ их дальнейшей организации в структуры более высокого порядка.
• Функциональный анализ нуклеосом, имеющий целью связать события, происходящие при активации хроматина и его транскрипции, со структурными изменениями.
К числу первоочередных вопросов, требующих ответа, относятся вопросы строения 30-нм фибриллы хроматина в контексте структуры нуклеосом и различных составляющих ее белков, а также связанные с механизмами активации и инактивации транскрипции через модификацию нуклеосом.
В 1970-е гг. основными репрессорами транскрипции считались гистоны, которые необходимо было нейтрализовать для того, чтобы ген получил возможность экспрессироваться. Однако предположения о том, каким образом этого можно достигнуть, не выходили за рамки представления об их удалении из состава хроматина с тем, чтобы обеспечить доступ активаторов к ДНК. С учетом состояния наших сегодняшних знаний мы можем считать, что находимся на пути познания взаимосвязи между функционированием хроматина и его строением.
Достигнут впечатляющий прогресс в исследовании роли ацетилирования и деацетилирования, а также других ферментативных модификаций гистонов, и мы начинаем понимать, как они изменяют локальную структуру хроматина и обеспечивают активацию промотора. Мы близки к объяснению механизмов активации промотора на основании этих структурных изменений. Рассматривая гистоны как репрессоры, мы начинаем понимать, каким образом при их взаимодействии с другими белками формируются области гетерохроматина, которые наследуются эпигенетически, и могут затрагивать как локальные, так и достаточно обширные области хроматина.
Мы ожидаем, что все эти события можно будет описать с точки зрения структурных изменений индивидуальных компонентов, т. е. в терминах молекулярных изменений отдельных гистонов и других белков хроматина.
Понимание всей иерархии высшего уровня организации хромосомы пока еще затруднено. Несмотря на колоссальный прогресс, достигнутый в области изучения генома, мы не до конца понимаем структурные особенности полос и промежутков в хромосомах, роль ГЦ-богатых последовательностей ДНК и т. д. Мы пока не можем связать эти характеристики с ДНК-белковыми взаимодействиями.
Даже несмотря на установление структуры 30-нм фибриллы хроматина, остается еще много неизвестного в отношении структур более высокого порядка. Такие существенные детали строения хромосомы как центромеры и теломеры немало способствовали пониманию молекулярной организации ее компонентов, хотя мы пока не в полной мере представляем себе роль последовательностей ДНК в организации центромерной области хромосом высших эукариот.
Так же как и в других областях биологии, возможность связать детали структуры с ее функциональными изменениями обеспечивает более высокий уровень познания процесса.
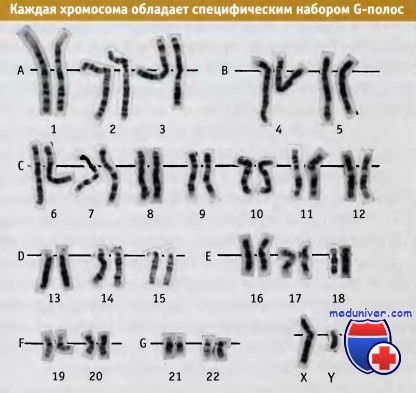
G-полосы образуют характерную поперечную исчерченность каждой хромосомы.
Генетический материал всех организмов и вирусов находится в форме плотно упакованного нуклеопротеина. В клетках эукариот транскрипционно активный хроматин находится в области эухроматина, который составляет большую часть интерфазного хроматина. Участки гетерохроматина упакованы в 3-10 раз более компактно и не проявляют транскрипционной активности.
В момент деления, когда становятся различимы индивидуальные хромосомы, весь хроматин приобретает плотную упаковку. О существовании ультраструктуры хромосом свидетельствует образование G-полос после их прокрашивания красителем Гимза. Эти полосы представляют собой очень большие области (порядка 107 пн), с помощью которых можно устанавливать места транслокации отдельных фрагментов или другие изменения структуры хромосом.
У эукариот интерфазный хроматин и метафазные хромосомы организованы в большие петлеобразные структуры. Каждая петля может представлять собой независимый суперспирализованный домен. Основания петлеобразных структур присоединяются к метафазно-му каркасу хромосомы или к ядерному матриксу по местам специфических последовательностей ДНК.
В области центромеры находится кинетохор, который обеспечивает присоединение хромосомы к митотическому веретену. Обычно центромера окружена гетерохроматином. Центромерные последовательности ДНК идентифицированы только для дрожжей S. cerevisiае, у которых они состоят из коротких консервативных элементов — CDE-I и CDE-III, связывающих комплексы CBF1 и CBF3 соответственно, и из протяженной АТ-богатой области, которая называется CDE-II. Эта область связывает Cse4p, образуя в хроматине специализированную структуру. Еще одна группа белков, которая связываются с этими комплексами, обеспечивают связь с микротрубочками.
Теломеры обеспечивают стабильность концов хромосомы. Почти все известные теломеры содержат множественные повторы, у которых одна цепь имеет состав Cn(A/T)m, n > 1 и m = 1-4. Вторая цепь, Cn(T/A)m содержит выступающий конец, который служит матрицей для добавления оснований в определенном порядке. Фермент теломераза представляет собой рибонуклеопротеиновый комплекс, в котором РНК представляет собой матрицу для синтеза цепи, богатой гуанином. Благодаря этому решается проблема репликации на самом конце дуплексной структуры. Теломера стабилизирует концы хромосом, поскольку выдающийся одноцепочечный участок Cn(T/A)m удаляет гомологичные участки в ранних повторяющиеся единицах теломеры, образуя петлю, и таким образом, свободные концы не возникают.
Хромосомы типа ламповых щеток у амфибий и политенные хромосомы насекомых характеризуются крайне релаксированной структурой, и коэффициент их упаковки составляет < 100. Политенные хромосомы D. melanogaster разделены на 5000 полос, размер которых варьирует на порядок и в среднем достигает 25 пн. Транскрипционно-активные области можно визуализировать даже в более релаксированных («пуффированных») структурах, у которых фибриллы хроматина распространяются от оси хромосомы. Это напоминает изменения, происходящие при транскрипции последовательностей эухроматина.
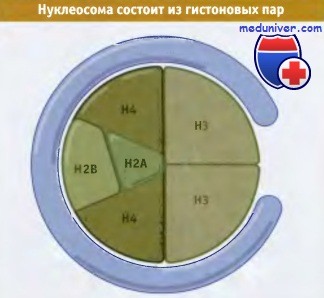
У эукариот хроматин состоит из нуклеосом. Нуклеосома содержит участок ДНК определенной длины, обычно -200 пн, обернутый вокруг октамера, состоящего из двух копий каждого гистона Н2А, Н2В, Н3 и Н4. С каждой нуклеосомой связана одна молекула гистона Н1. Почти вся геномная ДНК организована в нуклеосомы. Обработка нуклеазой микрококков показывает, что ДНК, упакованная в каждую нуклеосому, относится к одной из двух областей. ДНК линкерной области быстро переваривается эндонуклеазой, в то время как входящая в коровую область, размером 146 пн, более устойчива к действию фермента. Гистоны Н3 и Н4 относятся к числу наиболее консервативных, и тетрамер Н32-Н42 определяет размеры нуклеосомной частицы.
Гистоны Н2А и Н2В организованы в виде двух димеров Н2А-Н2В. Октамеры собираются при последовательном добавлении двух димеров Н2А-Н2В к ядру Н32-Н42.
В симметричной модели нуклеосомы тетрамер гистонов Н32-Н34 обладает формой ядра.
Один димер Н2А-Н2В виден сверху, второй находится снизу.
Нуклеосомы организованы в фибриллу диаметром 30-нм, которая содержит шесть нуклеосом на виток и характеризуется коэффициентом упаковки 40. При удалении гистона Н1 фибрилла раскручивается в 10-нм нить, состоящую из цепи нуклеосом. По-видимому, 30-нм фибрилла состоит из 10-нм нити, закрученной в соленоид. Эта фибрилла является основным компонентом эу- и гетерохроматина; в дальнейшей организации фибриллы в хроматин и хромосомы принимают участие негистоновые белки.
Существуют два пути сборки нуклеосом. Путь, связанный с репликацией, характеризуется активацией фактора сборки нуклеосом, CAF-1, которая происходит под действием процессивной реплисомной субъединицы — PCNA. Фактор CAF-1 обеспечивает фиксацию тетрамера Н32-Н42 на дочерних дуплексах, образующихся при репликации. Тетрамеры могут образовываться или при разрушении существующих нуклеосом репликативной вилкой, или в результате сборки из вновь синтезированных гистонов. Таким же путем образуются димеры Н2А-Н2В, которые затем добавляются к тетрамеру Н32-Н42, необходимому для построения нуклеосомы.
Поскольку тетрамеры Н32-Н42 и димеры Н2А-Н2В собираются случайным образом, новые нуклеосомы могут содержать как предсуществующие, так и новосинтезированные гистоны.
В ходе транскрипции РНК-полимераза удаляет октамеры гистонов. После прохождения полимеразы по ДНК нуклеосомы образуются вновь. Это наблюдается, когда транскрипция не носит столь интенсивный характер, как, например, в случае рДНК, когда они удаляются полностью. Образование нуклеосом по пути, независимому от репликации, происходит при возмещении октамеров гистонов, которые были удалены при транскрипции. При этом вместо гистона Н3 используется его вариант НЗ.З. По аналогичному пути происходит сборка нуклеосом на центромерных участках ДНК после окончания репликации, и также используется вариант Н3 гистона.
Ацетилирование гистонов происходит как при репликации, так и при транскрипции и необходимо для образования менее компактной структуры хроматина Некоторые белки, способствующие связыванию факторов транскрипции с ее основным аппаратом, проявляют ацетилазную активность по отношению к гистонам. Напротив, репрессоры часто ассоциированы с деацетилазной активностью. Ферменты модификации обычно проявляют специфичность по отношению к определенным аминокислотам в определенных гистонах. Наиболее общие сайты модификации локализованы на N-терминальных концах гистонов Н3 и Н4, которые выступают из нуклеосом между витками ДНК. Комплексы активации (или репрессии) обычно достигают больших размеров и часто характеризуются наличием нескольких ферментативных активностей, участвующих в различных модификациях компонентов хроматина.
Образование гетерохроматина происходит с участием белков, которые связываются со специфическими областями хромосом (таких, как теломеры) и взаимодействуют с гистонами. Образование неактивной структуры может распространяться вдоль фибриллы хроматина, начиная от центра инициации. Аналогичные события происходят в молчащих локусах типа спаривания у дрожжей.
Гетерохроматин начинает формироваться на определенных местах, и затем области распространяются на расстояние, величина которого не поддается точному определению. Когда гетерохроматин окончательно сформировался, характер распределения его областей наследуется в ходе протекания нескольких последовательных актов клеточного деления. Это служит примером эпигенетической наследственности, при которой две одинаковые последовательнсти ДНК могут быть связаны с различными белковыми структурами и поэтому обладают различной способностью к экспрессии. Таким образом объясняется наличие эффекта положения мозаичного типа у Drosophila.
Модификации «хвостов» гистонов запускают процесс реорганизации хроматина. Ацетилирование обычно связано с активацией генов. Ацетилазы гистонов присутствуют в комплексах активации, а деацетилазы — в комплексах инактивации. Метилирование гистонов сопряжено с инактивацией генов. Некоторые модификации гистонов могут проявлять синергический эффект.
Причина существования неактивного хроматина в теломерной области и в локусах типа спаривания у дрожжей связана с взаимодействием некоторых белков с N-терминальными «хвостами» гистонов Н3 и Н4. Образование неактивного комплекса может инициироваться связыванием одного белка на специфической последовательности ДНК; затем другие компоненты могут кооперативно связываться вдоль хромосомы.
Инактивация одной Х-хромосомы у женских особей млекопитающих (у плацентарных) происходит случайным образом. Для подсчета числа Х-хромосом необходимым и достаточным является локус Xic. Правило n-1 обеспечивает инактивацию всех Х-хромосом, кроме одной. Локус Xic содержит ген Xist, кодирующий РНК, которая экспрессируется только в неактивной Х-хромосоме. Стабилизация этой РНК представляет собой механизм, посредством которого одна Х-хромосома поддерживается в неактивном состоянии.