• Гетерохроматин формируется на специфических последовательностях, и неактивная структура распространяется по фибрилле хроматина
• Гены в области гетерохроматина находятся в неактивном состоянии
• Поскольку длина неактивного региона варьирует от клетки к клетке, инактивация генов в этой области обусловливает эффект положения мозаичного типа
• Аналогичные эффекты распространения проявляются на теломерах и на молчащих кассетах, контролирующих тип спаривания у дрожжей
Интерфазное ядро содержит как эу-, так и гетерохроматин. Степень конденсации гетерохроматина почти такая же, как в митотических хромосомах. Гетерохроматин не проявляет активности. В интерфазе он остается в конденсированном состоянии, транскрипция в нем подавлена, репликация приходится на позднюю S-фазу, и чаще всего он локализован на периферии ядра. Гетерохроматин центромеры обычно содержит сателлитную ДНК.
Однако образование гетерохроматина не зависит от первичной структуры ДНК. Когда ген перемещается за счет хромосомной транслокации или трансфекции и интегрируется в другое положение, ближе к гетерохроматину, в результате попадания на новое место он может инактивироваться. При этом предполагается, что он приобретает черты гетерохроматинового гена.
Мозаичный эффект положения, проявляющийся в изменении цвета глаз,
возникает, когда белый ген интегрируется рядом с гетерохроматиновым участком.
Клетки, в которых белый ген неактивен, образуют бляшки белоглазости, а клетки с активным белым геном образуют красные бляшки.
Выраженность эффекта определяется близостью интегрированного гена к гетерохроматиновой области.
Такая инактивация является результатом эпигенетического эффекта. Для отдельных клеток эукариот он может проявляться по-разному и приводить к эффекту положения мозаичного типа (PEV), при котором генетически идентичные клетки обладают различным фенотипом. Этот эффект хорошо изучен у Drosophila. иллюстрирует эффект положения на примере окраски глаза дрозофилы, некоторые фасетки которого не имеют окраски, в то время как другие окрашены в красный цвет.
Это объясняется тем, что белая окраска вызвана геном, который в некоторых клетках инактивировался под действием примыкающего гетерохроматина, в то время как в других остался активным.
На рисунке ниже представлено схематическое объяснение этого эффекта. Инактивация распространяется на разное расстояние от гетерохроматина в примыкающие области. В одних клетках она заходит достаточно далеко для того, чтобы инактивировать соседний ген, в других клетках этого не наблюдается. Это происходит в определенный момент эмбрионального развития, после чего состояние гена передается всем клеткам-потомкам. Потомки, произошедшие от клеток с инактивированным геном, образуют фасетки с фенотипом, отражающим выпадение признака (в данном случае неокрашенные, т. е. белого цвета).
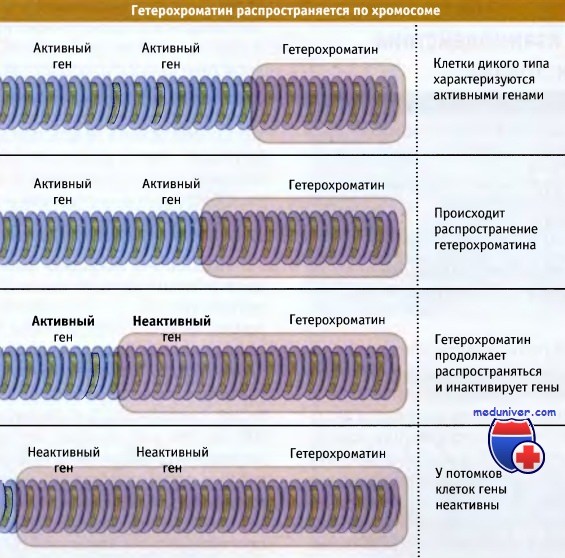
Чем ближе располагается ген к гетерохроматиновой области, тем выше вероятность того, что он будет инактивирован. Это позволяет предполагать, что образование гетерохроматина представляет собой двухэтапный процесс: вначале на специфической последовательности происходит нуклеация, а затем неактивное состояние распространяется по фибрилле хроматина.
Расстояние, на которое оно распространяется, точно не известно; скорее всего, процесс имеет стохастическую природу, и на него влияют такие параметры, как количество белковых компонентов, играющих ограничительную роль. К числу факторов, влияющих на процесс распространения, относится активация промоторов в определенной области; активный промотор может ингибировать распространение гетерохроматина.
Гены, расположенные поблизости от гетерохроматиновых областей, находятся в неактивном состоянии и поэтому в большинстве клеток неактивны. Эта модель предполагает, что границы гетерохроматиновой области могут меняться в зависимости от присутствия необходимых белков.
У дрожжей известен т. н. эффект теломерного молчания, аналогичный эффекту положения мозаичного типа у Drosophila: гены, локализованные в области тело-меры, в различной степени инактивированы. Это объясняется эффектом распространения гетерохроматина от теломеры.
Для дрожжей характерна еще одна форма эффекта молчания. Тип спаривания у них определяется активностью одного локуса (МАТ), однако в геноме содержатся две копии таких последовательностей (HML и HMR), которые находятся в неактивной конфигурации. Молчащие локусы HML и HMR имеют много общего с гетерохроматином, и их можно рассматривать как его небольшие области.
Распространение участка гетерохроматина приводит к инактивации генов.
Вероятность инактивации гена определяется его расстоянием от гетерохроматиновой области.