• Белки взаимодействуют между собой через небольшие консервативные домены
• Для передачи сигнала необходимы взаимодействующие модульные домены
• Адаптеры состоят исключительно из доменов или представляют собой связывающие последовательности
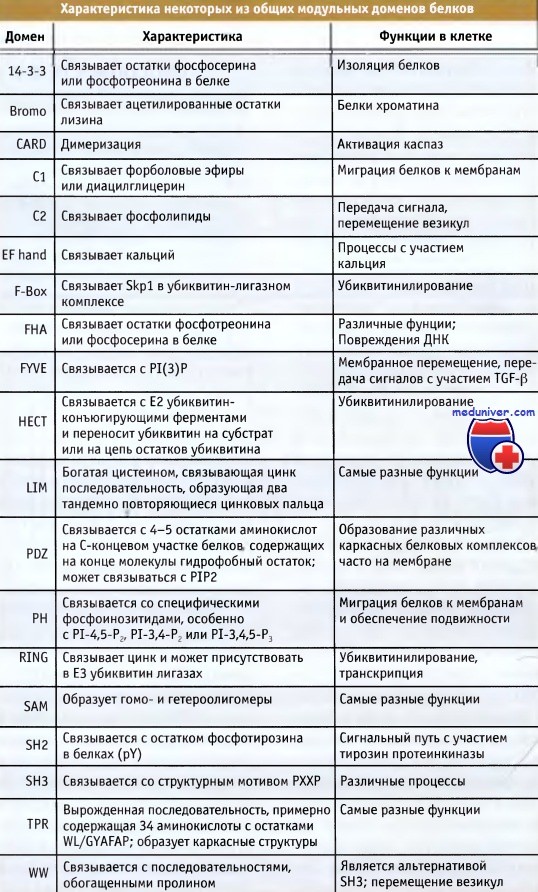
Модульные домены белок-белковых взаимодействий или связывающие белок последовательности присутствуют во многих сигнальных белках и обеспечивают их связывание с другими молекулами, включая белки, липиды и нуклеиновые кислоты. Некоторые из этих доменов перечислены на рисунке ниже. В противоположность каркасным структурам, которые связывают определенные белки достаточно селективно, во взаимодействии с модульными доменами обычно участвует не одна конкретная молекула, а группа мишеней, разделяющих общие структурные свойства.
Впервые модульные домены взаимодействий, участвующие в передаче сигнала, были обнаружены в тирозинкиназе, представляющей собой протоонкоген Src, которая содержит домен тирозинкиназы и два домена, называющиеся гомологичные Src (SH) домены 2 и 3. Первоначально модульные домены SH2 и SH3 были идентифицированы при сравнении структуры Src со структурой двух других тирозинкиназ, Fps и Abl. Один или оба этих домена присутствуют во многих белках и оба участвуют в белок-белковых взаимодействиях.
Домены SH3, которые содержат примерно 50 остатков аминокислот, связываются со специфическими короткими последовательностями, обогащенными пролином. Домены SH3 и богатые пролином последовательности присутствуют во многих белках цитоскелета, входящих в состав комплексов фокальной адгезии. Возможно, что такие последовательности могут направлять белки с доменами SH3 к этим специфическим клеточным сайтам. В отличие от сайтов SH2, связывающих фосфотирозин, сайты связывания с последовательностями богатыми пролином для доменов SH3 присутствуют в покоящихся и активированных клетках. Однако взаимодействие SH3 с пролином блокируется при фосфорилировании аминокислот в этой последовательности.
Домены SH2, которые содержат примерно 100 остатков аминокислот, связываются с белками, содержащими фосфорилированные остатки тирозина, например с цитоплазматической тирозинкиназой и с рецепторными тирозинкиназами. Таким образом, фосфорилирование Tyr регулирует свойства сайтов связывания SH2 и, тем самым, набор белок-белковых взаимодействий в зависимости от характера сигнала.
Для идентификации специфичности связывания доменов SH2 был использован элегантный метод. Вначале рекомбинантные SH2 домены инкубировали с клеточными лизатами, а затем с помощью аффинных методов их выделяли из инкубатов. Некоторые из белков, связанных с доменами SH2, взаимодействовали с антителами к фосфотирозину. С использованием этого и других методов было показано, что домены SH2 узнают последовательности, примыкающие к фосфотирозину, и для прочного связывания необходимо фосфорилирование тирозиновых остатков.
По мере идентификации природы взаимодействий, накапливаются данные о специфических последовательностях аминокислот, которые узнаются и связываются с модульными доменами. Такие данные также получаются в результате выполнения программы скрининга, с использованием библиотек кДНК и/или пептидов для оценки взаимодействий. На основании исследования специфичности последовательностей для индивидуальных доменов были идентифицированы консенсусные последовательности. Пользуясь этими последовательностями, можно предсказать, будет ли домен связываться с той или иной последовательностью в белке.
Адаптерные белки, которые не обладают ферментативной активностью, связывают сигнальные молекулы и располагают их таким образом, который позволяет им реагировать на внешние сигналы. Обычно адаптерные белки содержат два или более модульных доменов или связывающую последовательность. Адаптерные белки, в отличие от каркасных, обычно характеризуются многофункциональностью, поскольку их модульные домены и последовательности не обладают высокой специфичностью.
Адаптеры связываются с одним или несколькими другими сигнальными белками через домены белковых взаимодействий. Это приводит к группированию сигнальных белков или облегчает их дополнительные взаимодействия с другими белками.
Grb2 представляет собой прототип адаптерного белка, который был идентифицирован как белок, связывающийся с С-концевой областью EGF рецептора. Grb2 обладает одним SH2 и двумя SH3 доменами. Посредством SH3 домена он конститутивно связывается со специфическими обогащенными пролином участками белков, причем это связывание может блокироваться. Мишенью для Grb2 служит SOS, фактор обмена гуаниловых нуклеотидов, который активирует Ras, небольшой белок связывающий ГТФ, в ответ на сигнал, который подает EGF.
Посредством домена SH2, белок Grb2 связывает белки, фосфорилированные по тирозину, включая сами рецепторы, причем связывание носит сигнал-зависимый характер. Таким образом, фосфорилирование тирозина в этих рецепторах, в ответ на связывание ими лиганда, обеспечивает связывание их с Grb2, что, в свою очередь, приводит к перемещению SOS на рецептор, локализованный в мембране. Оказавшись в мембране, SOS может активировать свою мишень, Ras.
В таблице перечислены известные модульные домены белок-белковых взаимодействий, присутствующие во многих белках.
Белковые взаимодействия с участием этих доменов необходимы для контроля клеточных функций.
У отдельных прокариот обнаружено лишь несколько таких доменов.
По данным Паусон Лаб, Домены белковых взаимодействий.