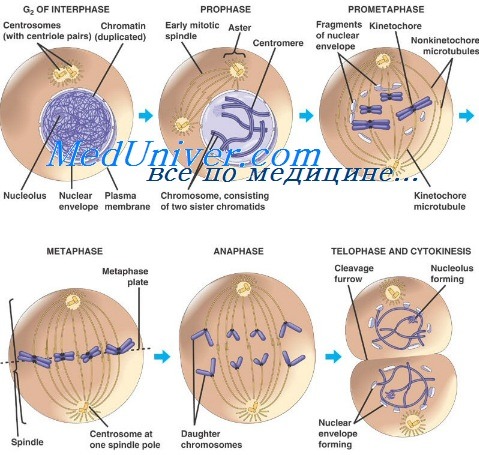
При митозе воспроизводится хромосомная организация клетки. Жизнь каждой дочерней клетки начинается с образования двух копий каждой хромосомы. Эти копии называют гомологами. Общее количество хромосом называют диплоидным набором, число хромосом в котором равно 2n. Во время интерфазы хромосомный материал в растущей клетке удваивается. В начале митоза каждая хромосома оказывается расщепленной по своей длине с образованием двух копий, называемых сестринскими хроматидами.
В этот момент число хромосом в клетке составляет 4n, они организованы в виде 2n пар сестринских хроматид. Процесс митоза состоит из 4 фаз: профазы, метафазы, анафазы и телофазы, заканчивающейся цитокинезом, когда делится сама клетка; при этом каждая дочерняя клетка получает тот же полный набор хромосом, который был в родительских клетках, а каждая хромосома в паре происходит от одной из родительских хромосом. Каждая фаза включает определенные движения центромеры — центральной сократительной области хромосомы.
Эти движения важны для разделения хромосомных пар между дочерними клетками и завершения процесса клеточного деления.
Непосредственно передмитозом происходит серия проверок в так называемых регуляторных точках (checkpoints) с целью репарации разрывов хромосомной двойной спирали или других повреждений ДНК. В нормальных условиях поврежденная ДНК восстанавливается, и процесс митоза завершается. Те клетки, где репарации ДИК не произошло, подвергаются апоптозу, или программируемой гибели. Такой механизм обеспечивает бесконечность деления клеток, если хромосомная ДНК интактна.
Результатом того, что клетке при делении удалось избежать контроля в одной из регуляторных точек, а клетки с разрывами двойной спирали или другими повреждениями ДНК делятся неконтролируемым образом, является карциногенез.
Важнейшими белками, участвующими в регуляции клеточного цикла, являются циклины и циклинзависимые киназы — CDK (суclin-dependent kinases). CDK — холоферментные комплексы, которые содержат регуляторные субъединицы И CDK-каталитичсскис субъединицы. Комплексы циклин-CDK регулируют 4 различные фазы клеточного цикла: Gap 1 (G1-фазу), фазу репликации ДНК (S-фазу), Gap 2 (G2-фазу) и собственно митоз (М-фазу). Контроль в точке рестрикции во время G1-фазы осуществляется при участии двух типов CDK: циклина D и циклина Е.
Циклины D-типа (D1, D2 и D3) во время ранней G1-фазы комбинаторно взаимодействуют С 2 каталитическими партнерами CDK4 и CDK6 с образованием по меньшей мере 6 холоферментов, экспрессия которых тканеспецифична. Циклин E образует комплекс со своим каталитическим партнером CDK2 и совместно с циклином D во время поздней G1-фазы завершает фосфорилирование белка-супрессора ретинобластомы (Kb). В результате клетка, минуя контроль регуляторной точки на участке G1-S, переходит в S-фазу.
Эндогенные ингибиторы комплексов циклин-CDK, называемые ингибиторами циклинзависимых киназ (CKI), экспрессируются во время G1-фазы и тормозят процессы фосфорилирования и активацию комплексов циклин-CDK, приводя к остановке G1 -фазы. Функция CKI состоит в том, чтобы не позволить клетке миновать регуляторную точку G1-фазы и ингибировать митоз, остановив тем самым рост клеток. На основании структуры самих ингибиторов и соответствующих CDK-мишеней CKI подразделяют на два семейства.
Белки семейства CIP/KIP (ингибиторы CDK) являются ингибиторами широкого спектра действия, они изменяют активность циклин D-, циклин В- и циклин А-завиеимых киназ. К этому семейству относятся белки р2l(Cip1), p27(Kipl) и p57(Kip2). Ike они в аминоконцевой области содержат характерную последовательность («мотив»), с которой связываются циклин н субстраты CDK. Белок р21 (Cipl) служит нижележащим эффектором фактора транскрипции и гена-супрессора опухоли p53 и обеспечивает репарацию повреждений ДНК и/или стимулирует апоптоз.
Белок p27(Kip1) является мощным ингибитором клеточной пролиферации в нормальных и патологически измененных тканях и выступает в роли важнейшего посредника при поражении тканей, воспалении и заживлении ран. Семейство белков INK4 (ингибитор CDK4) включает INK4A (p16), INK4B (р15), INK4C (р18) и INK4D (р19). Эти ингибиторы СОК содержат в своей молекуле анкириновые повторы, связываются только с CDK4 и CDK6, по не с другими CDK и специфически угнетают каталитические субъединицы CDK4 и СОКб, Белки семейства INK являются важнейшими регуляторами роста опухолей и играют большую роль в эволюционной биологии, однако их значение в патогенезе ССЗ не столь существенно.