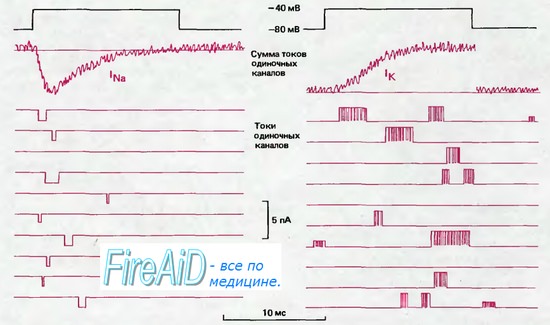
Рис. 2.12. Токи через натриевые (слева) и калиевые (справа) каналы (схематическое изображение). С помощью локальной фиксации потенциала производили сдвиг потенциала длительностью 14 мс от —80 до —40 мВ (черная линия); ниже показаны мембранные токи, зарегистрированные при нескольких таких последовательных сдвигах потенциала. Во время деполяризации токи одиночного канала могут возникать в любой момент, причем длительность их варьирует. При объединении многих записей токов в условиях синхронизации скачков потенциала получаются суммарные кривые токов, показанные вверху красным (lNa и lK). Временной ход lNa свидетельствует о том, что вероятность открывания Na+ -каналов наиболее высока вскоре после скачка потенциала, а примерно через 1 мс эти каналы открываются все реже и в конце концов инактивируются. Большая часть К+-каналов открывается с некоторой задержкой после скачка потенциала, затем средняя частота открываний остается на постоянном уровне в течение всего периода деполяризации.
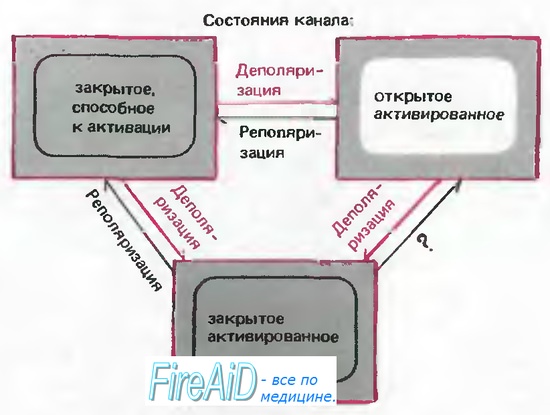
Рис. 2.13. Модель состояний Na+-каналов. «Закрытое, способное к активации» состояние при деполяризации может преобразовываться в «открытое активированное» или «закрытое инактивированное» состояние. Когда канал находится в «открытом активированном» состоянии, стойкая деполяризация способствует переходу в «закрытое инактивированное» состояние. Возвращение канала в «закрытое, способное к активации» состояние может происходить только в результате реполяризации. (Более реальная модель включает последовательно 3 «закрытых, способных к активации» и 4 «закрытых инак-тивированных» состояния [8].)
На рис. 2.12 справа схематически представлены токи одиночных К+-каналов, аналогично токам Na+-каналов (см. слева). Импульсы тока тоже имеют маленькую амплитуду (всего лишь +2 пА), а продолжительность открытого состояния канала варьирует вблизи среднего значения 5 мс. Однако в период открытого состояния К+-канал часто на короткое время закрывается, т. е. происходят быстрые осцилляции между открытым и закрытым состояниями. Такие «вспышки» открываний наблюдаются для многих типов каналов. В отличие от Na+-канала, К+-канал не инактивируется во время деполяризации; пока продолжается деполяризация, индивидуальные каналы непрерывно открываются и закрываются. В соответствии с этим, при суммации отведений получается кривая К+-тока, которая нарастает до стационарного уровня. Таким образом, описывая поведение токов К+-каналов с помощью модели, представленной на рис. 2.13, следует отметить, что инактивированное состояние в данном случае отсутствует, но наблюдаются два последовательных закрытых состояния, которые обеспечивают прерывистый характер вспышек [34] (см. Са2+-канал).
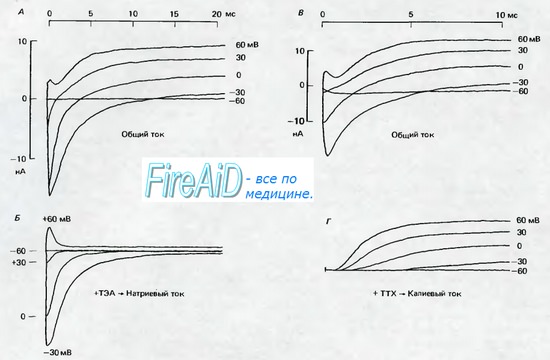
Рис. 2.6. А и В Мембранные токи в миелинизированных аксонах лягушки (перехваты Ранвье; 11 13 С) после ступенчатых сдвигов мембранного потенциала. Мембранный потенциал поддерживался с помощью фиксации потенциала на уровне потенциала покоя, равного —95 мВ; в момент времени 0 мс мембранный потенциал скачком поднимали до значений, которые указаны справа около записей тока, от —60 до +60 мВ. Сопровождающие скачок потенциала кратковременные емкостные токи вычитались, поэтому регистрируемые токи являются ионными токами. При —60 мВ скачок потенциала остается подпороговым и не вызывает изменений тока. По мере увеличения скачков потенциала сначала возникают отрицательные токи, которые с увеличением потенциала становятся положительными. Б. То же, что на рис. А, но на фоне блокады калиевых токов ТЭА (6 мМ), в результате чего токи почти полностью обеспечиваются ионами Na+. Полярность Na-токов меняется с отрицательной на положительную между значениями +30 и +60 мВ; по мере увеличения деполяризации продолжительность Na - токов уменьшается. Г. То же, что на рис. В. но на фоне блокады натриевых токов тетродотоксином (0,3 мкМ), так что записи соответствуют калиевым токам. При деполяризации К+-токи нарастают медленнее, чем Na - токи, и продолжаются в течение всего периода деполяризации (по [3] с изменениями)
Рис. 2.12 отражает поведение К+-каналов, типичное для нервных волокон: задержанное нарастание суммарного тока при деполяризации, заметное повышение проводимости во время деполяризации от потенциала покоя и отсутствие инактивации (ср. рис. 2.6). Обнаружено по крайней мере пять других типов К+-каналов. Они различаются, например, соотношением между открыванием канала и потенциалом мембраны, характеристиками инактивации или же зависимостью не только от деполяризации, но и от внутриклеточной концентрации Са2+. Эти типы К+-каналов обнаружены в клетках различных типов или частях клетки и присутствуют либо по отдельности, либо в виде определенных сочетаний. Именно разнообразие К+-ка-налов обусловливает вариации формы потенциалов действия, а также различную скорость реполяриза-ции и особенности следовых потенциалов (см. рис. 2.4). Существует яркий контраст между многообразием К+-каналов и одновременно Nа+-каналов, которые в возбудимых клетках животных всех типов быстро активируются деполяризацией, а затем быстро инактивируются.
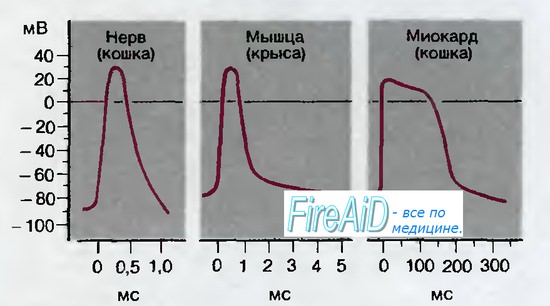
Рис. 2.4. Схематическое изображение потенциалов действия в различных тканях млекопитающих. Ордината: амплитуда внутриклеточного мембранного потенциала; абсцисса: время после начала потенциала действия. Временная шкала для каждого потенциала действия различна