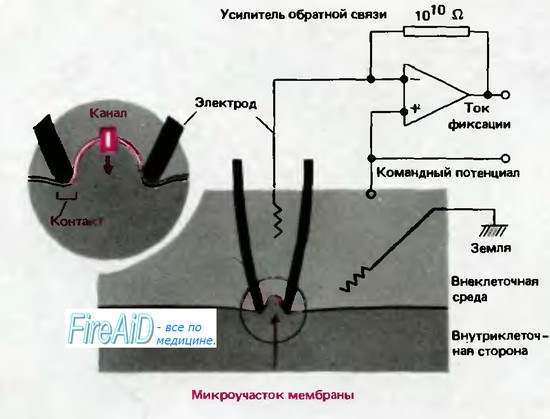
Рис. 2.11. Схема локальной фиксации мембранного потенциала («пэтч-кламп»). Изображен продольный срез через регистрирующую микропипетку (обозначена черным цветом) с диаметром контактирующего с мембраной кончика ~1 мкм. Если кончик электрода абсолютно чист и поверхность клетки освобождена от волокон соединительной ткани, то при подаче через пипетку отрицательного давления образуется тесный контакт, который создает электрическую изоляцию каналов находящегося в кончике пипетки микроучастка мембраны от остальной мембраны клетки (вставка). Таким способом можно регистрировать токи каналов с помощью усилителя обратной связи, соединенного с раствором электролита в пипетке (по [12, 24] с изменениями)
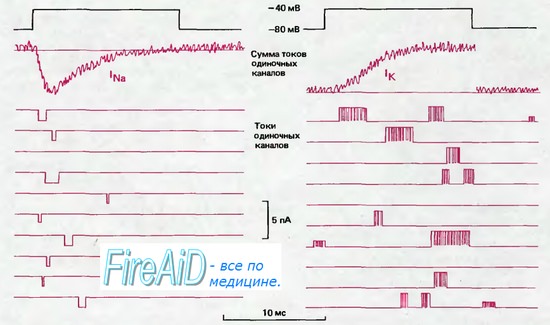
Токи через микроучасток мембраны, процедура регистрации которых показана на рис. 2.11, схематически представлены на рис. 2.12. Слева приведены 10 записей Na+-токa, при каждой из которых мембрана была деполяризована на период 14 мс. В каждом случае наблюдается только единственный короткий импульс тока с амплитудой —1,6 пА; это ток, протекающий через одиночную белковую молекулу Na+-канала. Длительность импульсов тока, которая соответствует времени открытого состояния канала, значительно варьирует около среднего значения 0,7 мс. Моменты открывания также варьируют, но при суммировании многих одиночных отведений получается результирующий временной ход тока, который на рис. 2.12 вверху слева изображен под записью скачка потенциала. Судя по записи временного хода тока, вероятность открывания канала резко возрастает при деполяризации, достигает максимума через 1,5 мс, затем снижается и становится минимальной через 10 мс после скачка деполяризации. Такое уменьшение вероятности открывания канала после деполяризации соответствует инактивации суммарного Na+-тока [8, 31].
Рис. 2.12. Токи через натриевые (слева) и калиевые (справа) каналы (схематическое изображение). С помощью локальной фиксации потенциала производили сдвиг потенциала длительностью 14 мс от —80 до —40 мВ (черная линия); ниже показаны мембранные токи, зарегистрированные при нескольких таких последовательных сдвигах потенциала. Во время деполяризации токи одиночного канала могут возникать в любой момент, причем длительность их варьирует. При объединении многих записей токов в условиях синхронизации скачков потенциала получаются суммарные кривые токов, показанные вверху красным (lNa и lK). Временной ход lNa свидетельствует о том, что вероятность открывания Na+ -каналов наиболее высока вскоре после скачка потенциала, а примерно через 1 мс эти каналы открываются все реже и в конце концов инактивируются. Большая часть К+-каналов открывается с некоторой задержкой после скачка потенциала, затем средняя частота открываний остается на постоянном уровне в течение всего периода деполяризации.
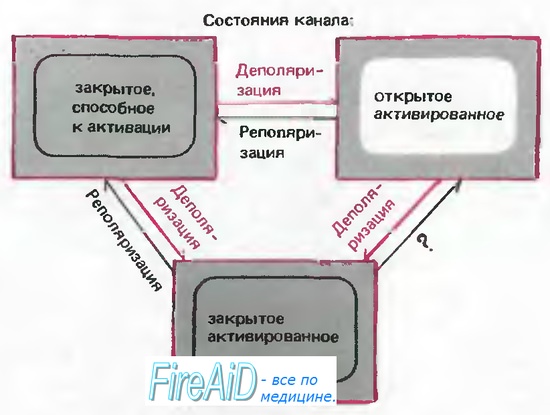
Рис. 2.13. Модель состояний Na+-каналов. «Закрытое, способное к активации» состояние при деполяризации может преобразовываться в «открытое активированное» или «закрытое инактивированное» состояние. Когда канал находится в «открытом активированном» состоянии, стойкая деполяризация способствует переходу в «закрытое инактивированное» состояние. Возвращение канала в «закрытое, способное к активации» состояние может происходить только в результате реполяризации. (Более реальная модель включает последовательно 3 «закрытых, способных к активации» и 4 «закрытых инак-тивированных» состояния [8].)
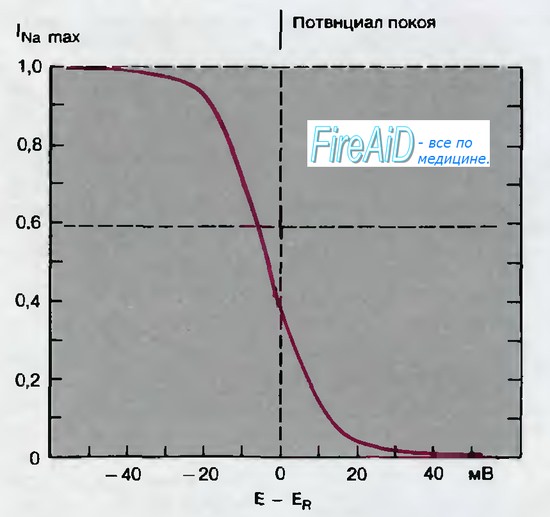
Отсюда следует, что открывание Na+-каналов при деполяризации не является строго детерминированным процессом; скорее происходит повышение вероятности открывания канала, а после того как он открылся, существует определенная вероятность, что он снова закроется. Таким «стохастическим» поведением обладают химические реакции, так что различные состояния канала-«закрытое, но способное к активации», «открытое» и «закрытое инактивированное» (неспособное к активации) можно связать между собой посредством постоянных скорости, как и в случае химических реакций. Простейшая модель поведения Na+-канала включает эти три состояния (рис. 2.13). Переход от закрытого и способного к активации в открытое состояние обеспечивается деполяризацией. Однако деполяризация ускоряет также и переход в инактивированное состояние, поэтому открытый канал подвергается быстрой инактивации и остается инактивированным, пока в результате ре- или гиперполяризации мембраны не вернется в закрытое, но способное к активации состояние. Равновесие между закрытым, но способным к активации и закрытым инактивированным состояниями тоже устанавливается посредством мембранного потенциала; это соотношение проявляется в виде зависимости от исходного потенциала способности Na+-токa к активации (рис. 2.8)
Рис. 2.8. Потенциал зависимая инактивация натриевой (Na)-системы. По оси абсцисс отложены величины отклонения мембранного потенциала от потенциала покоя ( — 60 мВ). От каждого из этих исходных значений потенциала мембрану деполяризовали до —16 мВ и по оси ординат откладывали отношения возникающих максимальных натриевых (Na+) -токов (lNamax) к величине lNamax, соответствующей полной активации натриевой (Na)-системы (по [15] с изменениями)