Промежуточная и долговременная регуляция потребления пищи. Терморегуляция и потребление пищи
а) Промежуточная и долговременная регуляция потребления пищи. Животные, голодавшие в течение длительного времени и затем получающие возможность съедать неограниченное количество пищи, потребляют ее существенно больше, чем регулярно питавшиеся. Напротив, животные, которых насильно кормили в течение нескольких недель, съедают очень мало пищи, когда им позволяют есть по собственному усмотрению. Таким образом, механизмы, регулирующие потребление пищи, находятся в зависимости от исходной обеспеченности питательными веществами.
б) Влияние концентрации глюкозы, аминокислот и жиров в крови на чувство голода и потребление пищи. Давно известно, что снижение концентрации глюкозы в крови вызывает ощущение голода. Это послужило поводом к формулированию так называемой глюкостатической теории регуляции голода и насыщения. Сходные результаты, но в связи с концентрацией аминокислот и продуктов расщепления жиров (кетокислота и некоторые жирные кислоты), привели к появлению аминостатической и липостатической теориям регуляции. Согласно этим концепциям, когда представленность в крови любого из трех видов питательных веществ снижается, это ведет к формированию пищевой мотивации, реализация которой в итоге возвращает концентрацию этих веществ в крови к нормальным характеристикам.
Нейрофизиологические исследования функции специфических областей мозга также поддерживают глюко-, амино-, липостатические концепции в связи со следующими наблюдениями:
(1) увеличение уровня глюкозы в крови приводит к увеличению частоты разрядов глюкорецепторных нейронов в вентромедиальном и паравентрикулярном ядрах центра насыщения гипоталамуса;
(2) аналогичное увеличение уровня глюкозы в крови одновременно снижает частоту разрядов в чувствительных к глюкозе нейронах в латеральных ядрах гипоталамического центра голода. Кроме того, некоторые аминокислоты и липиды влияют на частоту разрядов тех же нейронов или других, тесно с ними связанных.
в) Терморегуляция и потребление пищи. Если животные подвергаются воздействию холода, это ведет к увеличению потребления пищи. Когда они подвергаются воздействию жары, это создает тенденцию к снижению поступления энергии в организм. Это обусловлено взаимодействием гипоталамических центров терморегуляции и центров голода. Увеличение потребления пищи животными в условиях холода:
(1) увеличивает у них скорость метаболизма;
(2) обеспечивает увеличение жировых отложений, изолирующих тело от внешней среды.
Оба изменения препятствуют переохлаждению организма.
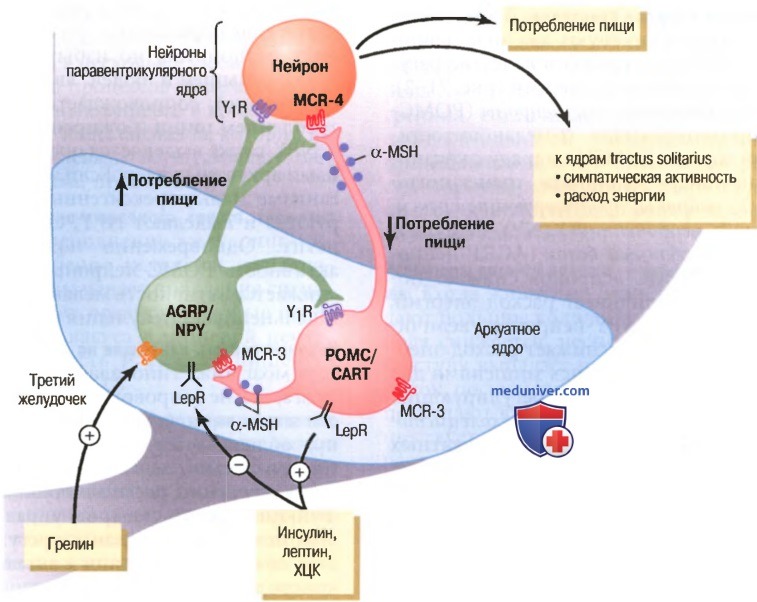
Регуляция энергетического баланса двумя типами нейронов аркуатных ядер:
(1) проопиомеланокортиновыми нейронами (РОМС), которые выделяют α-меланоцитостимулирующий гормон (α-MSH) и кокаин- и амфетамин-опосредованные транскрипты (CART), уменьшающими потребление пищи и увеличивающими расход энергии;
(2) нейронами, которые продуцируют меланин-опосредованный белок (AGRP) и нейропептид Y (NPY), увеличивающими потребление пищи и уменьшающими расход энергии; α-меланоцитостимулирующий гормон (α-MSH), выделяемый POMC-нейронами, стимулирует меланокортиновые рецепторы (MCR-3 и MCR-4) паравентрикулярных ядер, которые затем активируют нейрональный путь, проецирующийся на ядра солитарного тракта (tractus solitarius), и повышают симпатическую активность и расход энергии; AGRP действуют как антагонисты MCR-4.
Инсулин, лептин и холецистокинин, гормоны, ингибирующие AGRP- и NPY-нейроны и стимулирующие соседние РОМС- и CART-нейроны, уменьшают потребление пищи. Грелин (гормон, секретируемый желудком) активирует AGRP- и NPY-нейроны и стимулирует потребление пищи. LepR -рецепторы лептина, Y1R — рецепторы нейропептида Y1
г) Сигналы от жировой ткани регулируют потребление пищи по принципу обратной связи. Большую часть энергии организм запасает в виде жира, количество которого может значительно варьировать у разных индивидуумов. Чем регулируется этот энергетический резерв и почему существуют столь существенные индивидуальные различия?
Исследования последних лет дают основания предполагать, что гипоталамус узнает об энергетических запасах благодаря действию лептина, гормона пептидной природы, выделяемого адипоцитами. Если количество жировой ткани увеличивается (сигнал об избыточности жировых запасов), адипоциты продуцируют большое количество лептина, который поступает в кровь. Затем лептин переносится кровью к мозгу, проходит через гематоэнцефалический барьер путем облегченной диффузии и взаимодействует с рецепторами лептина, расположенными в разных областях гипоталамуса, особенно многочисленными на РОМС-нейронах аркуатных ядер и нейронах паравентрикулярных ядер.
Стимуляция рецепторов лептина в ядрах гипоталамуса инициирует многочисленные ответы, снижающие запасы жира, включая:
(1) снижение продукции гипоталамусом стимуляторов аппетита, таких как NPY и AGRP;
(2) активацию РОМС-нейронову высвобождающих MSH и активацию меланокортиновых рецепторов;
(3) продукцию гипоталамусом некоторых веществ, таких как кортикотропин-рилизинг гормон, который уменьшает потребление пищи;
(4) увеличение активности симпатической нервной системы (благодаря нервным связям гипоталамуса с сосудодвигательным центром), что увеличивает скорость метаболизма и расход энергии;
(5) снижение секреции инсулина β-клетками поджелудочной железы, ведущее к снижению энергетических запасов.
Таким образом, лептин может иметь важное значение как способ передачи сигнала от жировой ткани мозгу о том, что запасено достаточное количество энергии и дальнейшее потребление пищи не является необходимым. Мутации, возникшие у людей или мышей, за которые жировые клетки расплачиваются неспособностью продуцировать лептин, или мутации, являющиеся причиной дефектности рецепторов лептина, отмечены гиперфагией и патологическим ожирением.
Однако у людей с патологическим ожирением не отмечается дефицит продукции лептина, т.к. уровень лептина в плазме крови нарастает пропорционально развитию ожирения, поэтому некоторые физиологи предполагают, что ожирение может сочетаться с резистентностью к лептину. Это означает, что рецепторы лептина или рецепторные сигнальные пути, в норме активируемые лептином, могут быть дефектными при ожирении у людей, которые постоянно едят, несмотря на то, что уровень лептина в их крови очень высок.
Другим объяснением неспособности лептина предупреждать наращивание жировой ткани при ожирении в отдельных случаях может быть чрезмерное развитие систем, контролирующих пищевое поведение, направленное на предотвращение голодания, наряду с социальными и культурными факторами, которые могут формировать постоянное переедание даже в присутствии высокой концентрации лептина в крови.
д) Вывод. Информация о различных факторах, осуществляющих обратную связь в долговременной регуляции пищевого поведения, не является исчерпывающей, но можно утверждать: (1) если энергетические запасы в организме снижаются относительно нормальных показателей, то центр голода, расположенный в гипоталамусе и других отделах мозга, становится чрезвычайно активным, и индивидуум обнаруживает нарастающее чувство голода и высокую пищевую мотивацию для поиска пищи; (2) если энергетические запасы (главным образом запасы жира) достаточны, у индивидуума обычно отсутствует ощущение голода и формируется чувство насыщения.
Видео физиология терморегуляции - профессор, д.м.н. П.Е. Умрюхин