Гормоны регулирующие пищевое поведение. Нервная регуляция процесса еды
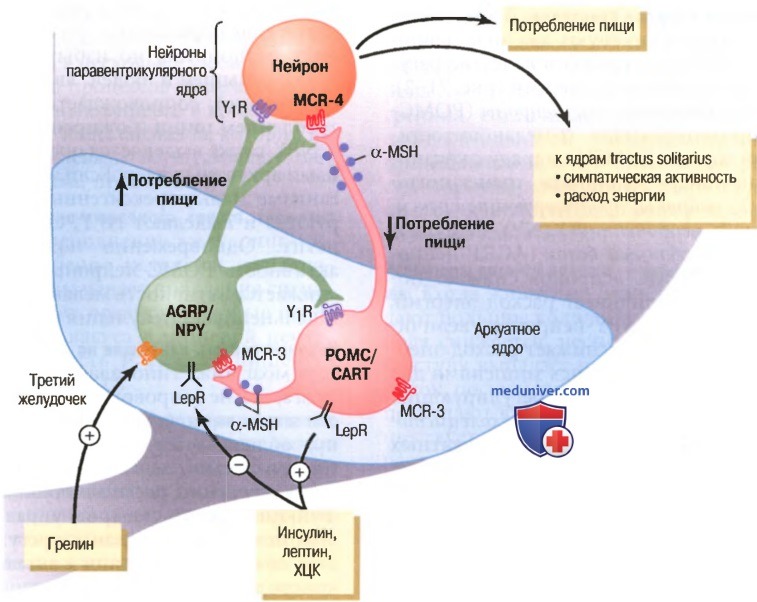
а) Нейроны и нейромедиаторы гипоталамуса, стимулирующие и подавляющие пищевое поведение. В аркуатном ядре гипоталамуса существуют два вида нейронов, которые особенно важны в качестве регуляторов аппетита и расхода энергии (для облегчения понимания просим вас изучить рисунок ниже):
(1) проопиомеланокортиновые нейроны (РОМС-нейроны), продуцирующие α-меланоцитостимулирующий гормон (α-MSH), наряду с кокаини амфетамин-опосредованными транскриптами (CART);
Регуляция энергетического баланса двумя типами нейронов аркуатных ядер:
(1) проопиомеланокортиновыми нейронами (РОМС), которые выделяют а-меланоцитостимулирующий гормон (a-MSH) и кокаин- и амфетамин-опосредованные транскрипты (CART), уменьшающими потребление пищи и увеличивающими расход энергии;
(2) нейронами, которые продуцируют меланин-опосредованный белок (AGRP) и нейропептид Y (NPY), увеличивающими потребление пищи и уменьшающими расход энергии; α-меланоцитостимулирующий гормон (α-MSH), выделяемый POMC-нейронами, стимулирует меланокортиновые рецепторы (MCR-3 и MCR-4) паравентрикулярных ядер, которые затем активируют нейрональный путь, проецирующийся на ядра солитарного тракта (tractus solitarius), и повышают симпатическую активность и расход энергии; AGRP действуют как антагонисты MCR-4.
Инсулин, лептин и холецистокинин, гормоны, ингибирующие AGRP- и NPY-нейроны и стимулирующие соседние РОМС- и CART-нейроны, уменьшают потребление пищи. Грелин (гормон, секретируемый желудком) активирует AGRP- и NPY-нейроны и стимулирует потребление пищи. LepR -рецепторы лептина, Y1R — рецепторы нейропептида Y1
Активация РОМС-нейронов снижает потребление пищи, увеличивает расход энергии, активация же AGRP- и NPY-нейронов увеличивает потребление пищи и снижает расход энергии. Данные нейроны оказались мишенями для действия некоторых гормонов, регулирующих аппетит, включая лептин, инсулин, холецистокинин и грелин. Фактически нейроны аркуатных ядер являются областью, где сходятся многочисленные центральные рефлекторные и периферические сигналы, регулирующие депонирование энергии.
РОМС-нейроны высвобождают α-MSH, действующий потом на меланокортиновые рецепторы (MCR), обнаруженные главным образом в нейронах паравентрикулярных ядер. Хотя существуют, по меньшей мере, пять подтипов мела-нокортиновых рецепторов, MCR-3 и MCR-4 особенно важны в регуляции пищевого поведения и энергетического баланса. Активация этих рецепторов снижает потребление пищи, увеличивая при этом расход энергии. Напротив, торможение этих рецепторов заметно увеличивает потребление пищи и снижает расход энергии.
Влияние активации MCR увеличивает расход энергии, по крайней мере частично, активацией нейрональных путей, идущих от паравентрикулярных ядер к ядрам одиночного пучка и повышающих активность симпатической нервной системы.
Гипоталамическая меланокортиновая система играет существенную роль в регуляции запасания энергии в организме, и нарушения в сигнальных меланокортиновых путях сочетаются с резко выраженным ожирением. Установлено, что мутации MCR-4 в 5-6% случаев являются причиной рано наступающего тяжелого ожирения у детей. Напротив, выраженная активация меланокортиновой системы снижает аппетит. Некоторые исследователи предполагают, что такая активация может быть причиной анорексии, встречающейся на фоне тяжелых инфекций и раковых опухолей.
AGRP, высвобождаемый из орексигенных нейронов гипоталамуса, является естественным антагонистом активаторов MCR-3 и MCR-4 и, возможно, увеличивает потребление пищи путем ингибирования влияний MSH, стимулирующего меланокортиновые рецепторы (для облегчения понимания просим вас изучить рисунок выше). Роль AGRP в регуляции потребления пищи при нормальных физиологических условиях не понятна, но избыточное образование AGRP у мышей и людей, являющееся следствием мутаций, сопровождается избыточным потреблением пищи и ожирением.
NPY также выделяется орексигенными нейронами аркуатных ядер. Если запасы энергии в организме малы, орексигенные нейроны активируются и выделяют NPY, стимулирующий аппетит. Одновременно подавляется залповая активность РОМС-нейронов, вследствие этого снижается активность меланокортиновых путей и дальнейшая стимуляция аппетита.
Нервные центры, влияющие на процесс еды. Если пересечь мозг ниже гипоталамуса, но выше среднего мозга, это не сопровождается утратой способности к поеданию пищи. Животное выделяет слюну, облизывается, жует, проглатывает пищу. Следовательно, механизмы акта еды организуются центрами, расположенными в стволе мозга. Функции прочих центров, управляющих пищевым поведением, связаны с регуляцией количества потребляемой пищи и активацией центров, контролирующих сам акт еды.
Нервные центры, расположенные выше гипоталамуса, также играют важную роль в регуляции пищевого поведения, особенно в управлении аппетитом. Эти центры включают амигдалу (миндалину) и префронтальную кору, обладающую тесными связями с гипоталамусом. Уместно напомнить, что в отдельной статье на сайте (просим вас пользоваться формой поиска выше) обсуждался вопрос о вкладе амигдалы в восприятие запахов. Разрушения амигдалы продемонстрировали, что некоторые ее области ответственны за стимуляцию пищевого поведения, в то время как другие ингибируют его. Важным эффектом двустороннего разрушения амигдалы является психическая «слепота» в выборе пищи. Другими словами, животные (а иногда и люди) утрачивают или частично утрачивают способность контролировать качество съедаемого, в частности отличать съедобное от несъедобного.