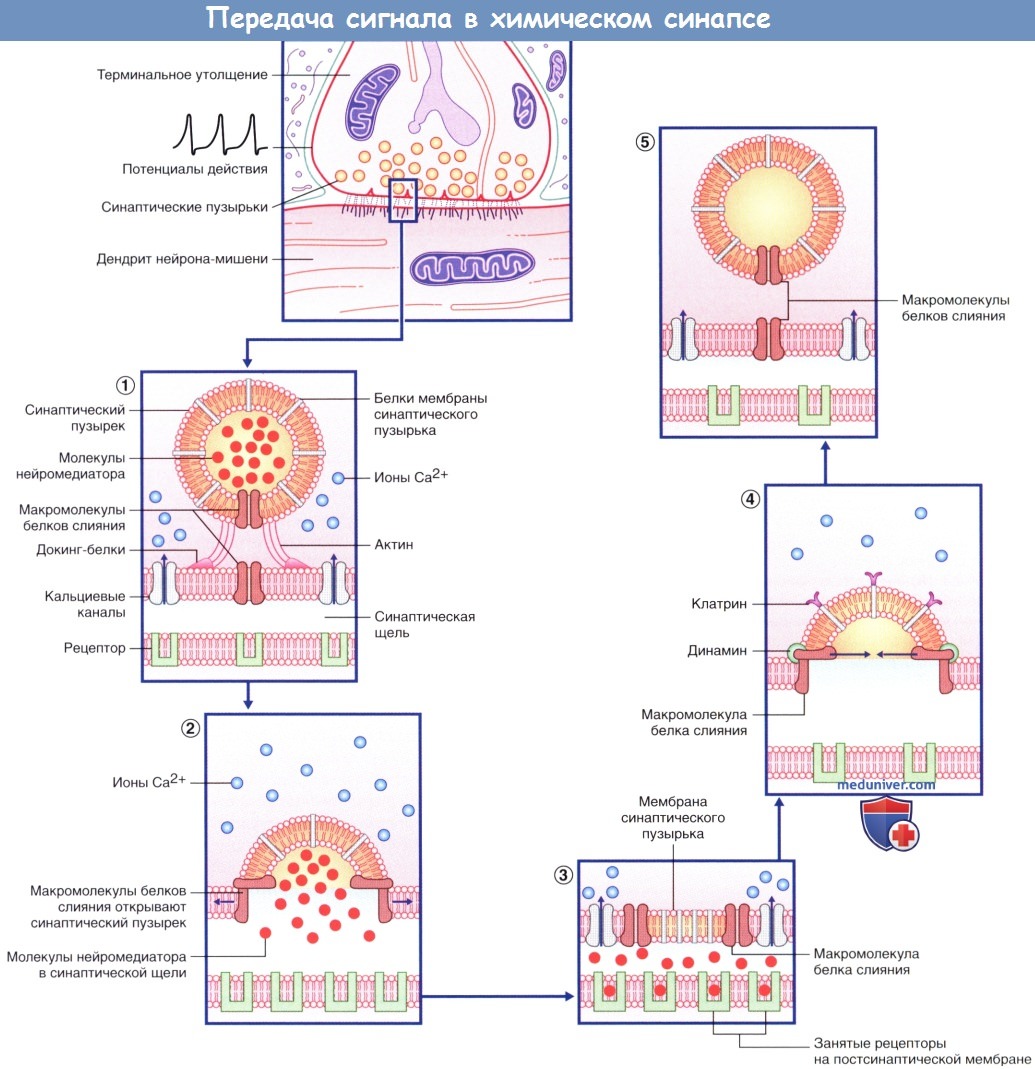
а) Высвобождение медиаторов. В состоянии покоя синаптические пузырьки накапливаются в активных зонах нервных окончаний и направляются к пресинаптическим уплотнениям при участии докинг-белков, один из которых — актин. При прохождении потенциала действия происходит открытие потенциалозависимых кальциевых (Са2+) каналов, расположенных в непосредственной близости к активной зоне пресинаптической мембраны, что приводит к быстрому заполнению этой области ионами Са2+. Ионы Са2+ связываются с определенными белками, расположенными в области скопления синаптических пузырьков и в активной зоне, в результате чего происходят слияние синаптических пузырьков с клеточной мембраной и высвобождение нейромедиаторов.
Процесс слияния синаптических пузырьков с клеточной мембраной начинается с образования плотного комплекса между везикулярными белками SNARE (v-SNARE) и белками пресинаптической мембраны (t-SNARES). Метаболические компоненты, необходимые для осуществления слияния синаптических пузырьков с мембраной клетки, расположены в активной зоне. Наиболее важный крупный мультидоменный белок RIM соединяется с ГТФ-связывающим белком синаптических пузырьков Rab3. Остальные белки обеспечивают слияние синаптических пузырьков с мембраной клетки и высвобождение нейромедиаторов. К синаптическим компонентам относят синаптические везикулярные белки (синаптотагмин, синаптобревин, синаптофизин и синапсины), везикуло-ассоциированные белки (амфифизин, динамин и СаМ-киназы), синаптические белки плазматической мембраны (синтаксины, нейрексины и SNAP-25), а также цитозольные белки (комплексины, различные виды белков SNAP и NSF-белки).
Многие из перечисленных синаптических белков выполняют специфические функции по обеспечению единства процессов возбуждения клетки и высвобождения нейромедиаторов, а также механизмов восстановления синаптических пузырьков, что лежит в основе синаптической передачи сигнала. Молекулярные процессы, обеспечивающие взаимосвязь возбуждения клетки и высвобождения нейромедиаторов, представляют собой сложные модели, осуществляемые различными механизмами. Белки сразу же формируют ответную реакцию на поступление ионов Са2+ в клетку, в связи с чем временной промежуток между входом ионов и высвобождением нейромедиаторов составляет менее 1 мс.
Для высвобождения некоторого количества нейромедиаторов из синаптических пузырьков небольшого размера [например, содержащих глутамат или у-аминомасляную кислоту (ГАМК)] достаточно единичных потенциалов действия. В это же время для запуска медленного (с задержкой 50 мс и выше) высвобождения медиаторов из гранулярных везикул большого размера, характерных для пептидергических нейронов, минимальная частота импульсов составляет 10 Гц. Таким образом, количество высвобождаемого медиатора не является постоянной величиной и может изменяться под влиянием внутренних и внешних факторов.
Последовательность процессов, происходящих в клетке после деполяризации пресинаптической мембраны.
(1) При открытии кальциевых каналов (Са2+) (указано стрелкой) за счет актиновых филаментов синаптические пузырьки приближаются к пресинаптической мембране.
Парные макромолекулы белков слияния (FPMS), расположенные на синаптическом пузырьке, совмещаются с соответствующими белками на пресинаптической мембране.
(2) Макромолекулы белков слияния разделяются (указано стрелками, направленными в разные стороны), тем самым обеспечивая выход молекул нейромедиатора в синаптическую щель.
(3) Мембрана синаптического пузырька встраивается в пресинаптическую мембрану, и нейромедиаторы активируют специфические рецепторы.
(4) Молекулы белка клатрина участвуют в перемещении мембраны синаптического пузырька внутрь клетки. Молекулы белка динамина (выделены зеленым цветом) участвуют в сближении пар макромолекул белков слияния (указано стрелками, направленными навстречу друг другу) и замыкают формирующийся синаптический пузырек.
(5) Синаптический пузырек не содержит медиатора и может быть использован повторно.
б) Связывание медиатора с рецептором клетки-мишени. Молекулы нейромедиатора связываются с молекулами белков-рецепторов на постсинаптической мембране. Выделяют ионотропные и метаботропные рецепторы. Каждая группа включает рецепторы, активация которых приводит к открытию ионных каналов, а также рецепторы, обеспечивающие закрытие этих каналов.
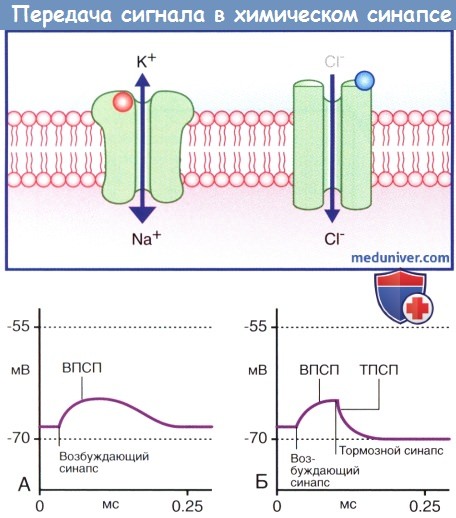
1. Ионотропные рецепторы. В каждой макромолекуле ионотропных рецепторов присутствует ионный канал. Медиатор связывается со специфическим рецептором в синаптической щели, вызывая ее конформационное преобразование, что приводит к открытию ионного канала. Считают, что ионотропные каналы являются медиаторозависимыми (или лиганд-зависимыми), что указывает на их способность связываться с молекулой медиатора или лекарственного вещества. После отсоединения молекулы нейромедиатора или его разрушения ионный канал, закрываясь, возвращается в исходное состояние.
Нейромедиатор вызывает открытие возбуждающего ионного канала, в результате чего происходят значительное поступление ионов натрия (Na+) в клетку и незначительный выход ионов калия (К+) из клетки. Эти процессы приводят к образованию возбуждающего постсинаптического потенциала (ВПСП), вызывающего деполяризацию мембраны. При открытии множества медиаторозависимых каналов происходит суммирование возбуждения и возникает деполяризация большей силы, которая может достигать пороговых значений и вызывать образование потенциала действия. Тормозной постсинаптический потенциал (ТПСП), следующий за ВПСП, вызывает гиперполяризацию мембраны до -70 мВ, что служит равновесным потенциалом для ионов хлора (Cl-). При открытии калиевых каналов, равновесный потенциал ионов которых составляет -80 мВ, происходит гиперполяризация большей силы.
Ионотропные рецепторы получили название «быстрых», поскольку они оказывают быстрое, но непродолжительное действие на ионные каналы.
(А) Медиаторозависимый возбуждающий ионотропный рецептор.
Присоединение нейромедиатора (в данном случае изображен глутамат, выделен красным цветом; возбуждающий синапс) приводит к открытию «смешанного» натрий-калиевого канала.
При большом поступлении ионов Na+ в клетку происходит незначительный выход ионов К+ из клетки, что приводит к деполяризации мембраны и возникновению возбуждающего постсинаптического потенциала (ВПСП).
(Б) Медиаторозависимый тормозной ионотропный рецептор. Последующее присоединение тормозного нейромедиатора (в данном случае изображен ГАМКA, выделен синим цветом; возбуждающий синапс) приводит к открытию хлорного ионного канала.
Ионы Cl- поступают внутрь клетки, и под действием тормозного постсинаптического потенциала (ТПСП) происходит гиперполяризация мембраны.
2. Метаботропные рецепторы. Метаботропные рецепторы получили свое название благодаря способности оказывать в цитоплазме нейрона многочисленные метаболические действия. Макромолекула рецептора представляет собой трансмембранный белок, в котором отсутствует ионный канал. Рецептор активируется посредством присоединения медиатора к его активному центру, в результате чего происходит конформационная перестройка белка, что, в свою очередь, активирует одну из закрепленных субъединиц (α- или β-субъединицу). После активации субъединица открепляется и направляется вдоль внутренней поверхности клеточной мембраны. Субъединицы являются G-белками за счет преимущественного связывания с гуанинтрифосфатом (ГТФ) или гуаниндифосфатом (ГДФ).
В большинстве случаев эти белки оказывают непрямое действие через систему вторичных посредников. Однако некоторые G-белки активируют ионные каналы напрямую. G-белок, оказывающий возбуждающее действие, называют Gs-белком, а белок, оказывающий тормозное действие,— Gi-белком. За счет многостадийного функционирования метаботропные рецепторы, как правило, являются «медленными»; после единичного стимула влияние мембранного канала может продолжаться сотни миллисекунд. Кроме того, образование вторичных посредников может привести к снижению способности нейрона к возбуждению.
В настоящее время выделяют три системы вторичных посредников.
1. Система циклического аденозинмонофосфата (цАМФ), осуществляющая фосфорилирование белков.
2. Инозитолфосфатная система, обеспечивающая высвобождение ионов Са2+ из эндоплазматических депо.
3. Система арахидоновой кислоты, за счет которой осуществляется образование метаболитов арахидоновой кислоты.
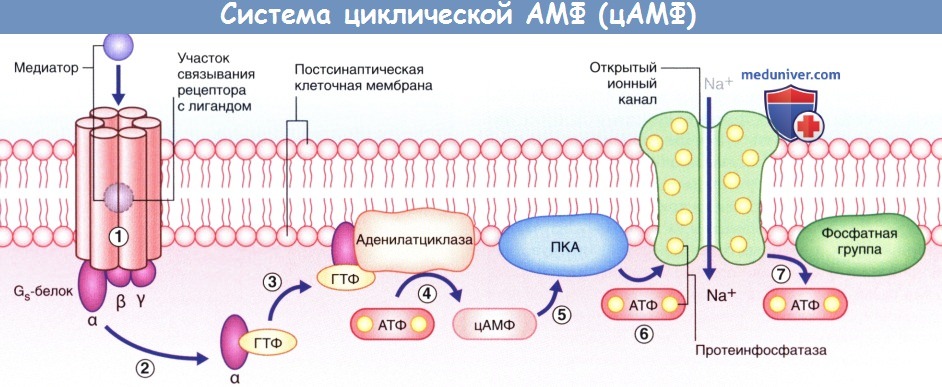
- Система цАМФ. Связывание нейромедиатора с рецептором приводит к отделению а-субъединицы от Gs-белка, за счет чего становится возможным присоединение ГТФ к освободившемуся белку, который, в свою очередь, способствует превращению аденозинтрифосфата (АТФ) в цАМФ под действием аденилатциклазы (рис. 8.4). Синтезированный цАМФ, расщепляясь, выполняет в клетке функцию вторичного посредника. Под воздействием цАМФ протеинкиназа А переносит фосфат-ионы от молекулы АТФ в ионный канал, что приводит к его открытию и поступлению ионов Na+ внутрь клетки, за счет чего происходит деполяризация нейрона-мишени. При инактивации Gs-белка под действием прикрепленного к мембране фермента протеинфосфатазы происходят выход фосфат-ионов из клетки и закрытие ионного канала.
Система циклической АМФ (цАМФ).
На рисунке изображена последовательность процессов, происходящих в клетке, начиная от Gs-белок-связанного рецептора к ионному каналу через образование цАМФ.
(1) Медиатор активирует макромолекулу рецептора.
(2) α-Субъединица Gs-белка отщепляется и связывается с гуанозинтрифосфатом (ГТФ).
(3) ГТФ связывается с аденилатциклазой.
(4) Под действием аденилатциклазы происходит синтез цАМФ из АТФ.
(5) цАМФ активирует протеинкиназу А (ПКА).
(6) ПКА перемещает фосфатные группы АТФ на натриевый (Na+) канал, за счет чего канал открывается, и ионы Na+-поступают в цитозоль, вызывая деполяризацию клетки.
(7) Вслед за инактивацией Gs-белка под действием фермента фосфотазы происходят дефосфорилирование ионного канала и его закрытие.
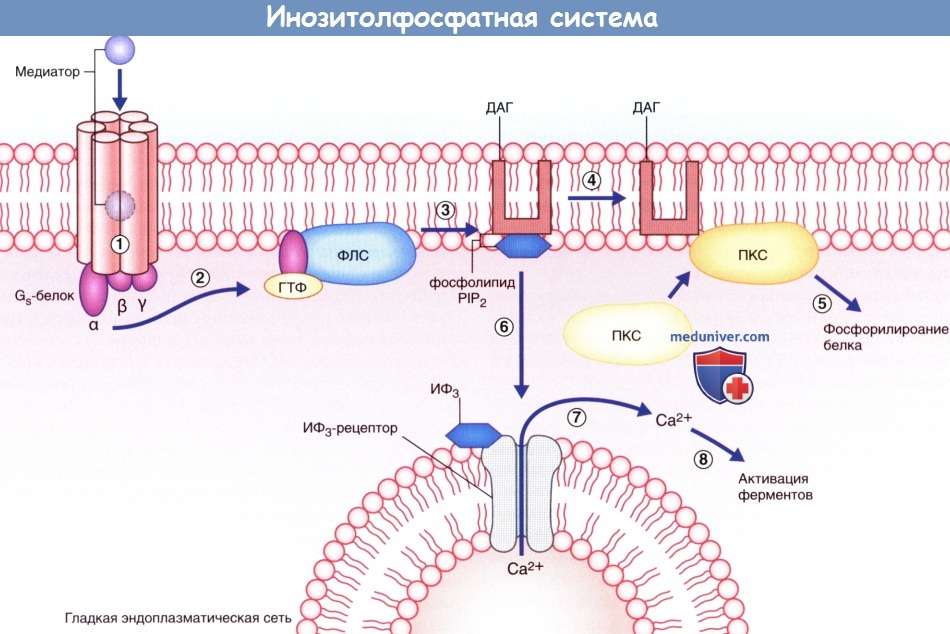
- Инозитолфосфатная система. Эффекторный фермент — фосфолипаза С — под влиянием другой активированной а-субъединицы G -белка расщепляет фосфолипид мембраны PIP, с образованием двух вторичных посредников; диацилглицерола (ДАГ) и инозитолтрифосфата (ИТФ). ДАГ активирует протеинкиназу С, которая запускает процесс фосфорилирования белков. ИТФ, проникая в цитозоль, открывает кальций-зависимые ионные каналы преимущественно вблизи мембран гладкой эндоплазматической сети. Ионы Са2+ запускают Са2+-зависимый каскад ферментов, осуществляющих открытие и/или закрытие ионных каналов, а также, вероятно, изменяющих экспрессию генов и синтез белков при прохождении через ядерную оболочку (см. далее — транскрипция генов).
Система арахидоновой кислоты. Описана далее (в связи с гистамином).
Инозитолфосфатная система.
Продемонстрирована двойная функция этой системы.
(1) Медиатор активирует макромолекулу рецептора.
(2) α-Субъединица выбелка отщепляется и связывается с гуанозинтрифосфатом (ГТФ), который обеспечивает ее связывание с фосфолипазой С (ФПС).
(3) ФПС перемещается вдоль клеточной мембраны и расщепляет фосфолипид РIР2 на диацил-глицерол (ДАГ) и инозитолтрифосфат (ИФ3).
(4) ДАГ вызывает перемещение протеинкиназы С (ПКС) к мембране клетки, в результате чего происходит (5) фосфорилирование некоторых белков, в том числе и белков ионных каналов.
(6) ИФ3 активирует кальциевые (Са2+) каналы гладкой эндоплазматической сети.
(7) Ионы Са2+-поступают в цитозоль. (8) Происходит активация Са2+-зависимых ферментов.
- Транскрипция генов. В настоящее время установлено, что рефлекторный ответ на повторяющиеся воздействия способен либо прогрессивно увеличиваться при формировании сенсибилизации, что в большинстве случае происходит под влиянием вредоносных стимулов, либо уменьшаться при формировании приспособления, что чаще встречается при воздействии безвредных стимулов. В ходе экспериментов на животных, изучались рефлекторные дуги, в которых были задействованы чувствительные, двигательные и ассоциативные нейроны, было показано, что для развития сенсибилизации характерно образование новых синаптических контактов между ассоциативными и двигательными нейронами; это сопровождается дополнительным синтезом и высвобождением медиаторов. Для развития приспособления, напротив, характерно снижение синтеза и высвобождение медиаторов. Данные процессы обусловлены изменением транскрипции генов.
Повторяющиеся вредоносные стимулы оказывают влияние на цАМФ, в результате чего происходит чрезмерная активация протеинкиназ, участвующих в фосфорилировании белков, регулирующих транскрипцию генов. При этом увеличивается синтез белков (в том числе и ферментов), необходимых для синтеза медиаторов и построения новых ионных каналов и цитоскелета синапса. Действие повторяющихся безвредных стимулов заключается только в снижении синтеза и высвобождения медиаторов.
Влияния транскрипции генов играют особенно значимую роль в процессе формирования долгосрочной памяти.
Видео физиология синапса и нерва - профессор, д.м.н. П.Е. Умрюхин