Наиболее распространенный возбуждающий медиатор головного и спинного мозга — аминокислота L-глутамат. Значимый пример возбуждающих нейронов, использующих глутамат в качестве медиатора,— все нейроны, идущие от коры полушарий к белому веществу мозга, независимо от их направления в других частях коры полушарий, ствола или спинного мозга. Глутамат синтезируется из α-кетоглутарата, который, кроме того, служит субстратом для образования ГАМК.
ГАМК — самый распространенный в спинном и головном мозге тормозной медиатор, участвующий в работе приблизительно трети всех синапсов нервной системы. Миллионы ГАМКергических нейронов образуют основную часть вещества хвостатого и чечевицеобразного ядер, их также встречают в большом количестве в околоводопроводном сером веществе, гипоталамусе и гиппокампе. Кроме того, ГАМК выполняет функцию медиатора в крупных клетках Пуркинье, которые являются единственными клетками, выходящими из коры мозжечка. Аксоны клеток Пуркинье спускаются к зубчатому и другим ядрам мозжечка. ГАМК синтезируется из глутамата под действием фермента глутаматдекарбоксилазы.
Третий аминокислотный нейромедиатор — глицин. Глицин участвует в синтезе белков всех тканей организма и представляет собой простейшую аминокислоту, синтезируемую из серина в процессе катаболизма глюкозы. Этот нейромедиатор оказывает тормозное действие преимущественно в синапсах ассоциативных нейронов ствола мозга и спинного мозга.
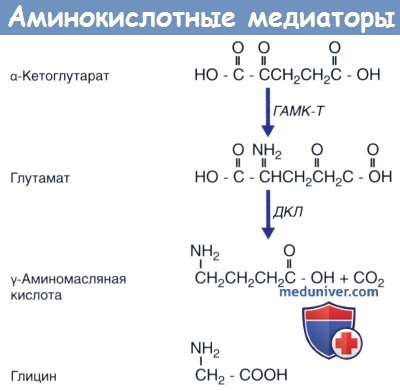
Три аминокислотных медиатора.
Глутамат синтезируется из а-кетоглутарата под действием фермента ГАМК-трансаминазы (ГАМК-Т);
γ-аминомасляная кислота (ГАМК) синтезируется из глутамата под действием декарбоксилазы глутаминовой кислоты (ДГК).
Глицин представляет собой простейшую аминокислоту.
а) Глутамат. Глутамат выполняет функцию нейромедиатора как в ионотропных, так и в метаботропных рецепторах. К ионотропным рецепторам относят АМРА-, каинат- и NMDA-рецепторы, которые получили свои названия благодаря активирующим их синтетическим агонистам: амино-метил-изоксазол-пропионовой кислоте, каинату и N-метил-D-аспартату, соответственно. Каинатные рецепторы редко встречаются изолированно; чаще всего они комбинируются с АМРА-рецепторами и входят в состав АМРА-каинатных (АМРА-К) рецепторов.
Ионотропные рецепторы глутамата. При активации АМРА-К-рецепторов на постсинаптической мембране происходят немедленное поступление большого количества ионов Na+ в клетку и выход небольшого количества ионов К+ из клетки, что приводит к формированию раннего компонента ВПСП нейрона-мишени, деполяризующего мембрану клетки-мишени от -65 мВ до -50 мВ. Этот процесс приводит к электростатическому «выталкиванию» катионов магния (Mg2+), которые в состоянии покоя «закрывают» ионный канал NMDA-рецептора. Ионы Na+ проходят через ионный канал, происходит формирование потенциала действия.
Важно отметить, что ионы Са2+ также проникают внутрь клетки и за счет продолжительного периода деполяризации, длительность которого достигает 500 мс от возникновения единичного потенциала действия, активируют Са2+-зависимые ферменты, способные изменять структуру клетки-мишени и даже количество ее синаптических контактов. Феномен синаптической пластичности в ответ на активацию рецепторов можно отчетливо проследить в экспериментальных исследованиях на культивируемых срезах гиппокампа крыс. Этот феномен считают основным механизмом развития кратковременной памяти. Например, анальгетик кетамин, блокирующий NMDA-каналы, помимо своего основного действия препятствует формированию памяти.
Характерная особенность многократно повторяющейся активации NMDA-рецепторов — долговременное потенцирование, проявляющееся возникновением ВПСП со значениями, превышающими нормальные показатели даже несколько дней спустя (см. далее — длительная депрессия).
Роль NMDA-рецепторов в развитии феномена глутаматной эксайтотоксичности подтверждена развитием ишемических инсультов у экспериментальных животных. Предполагают, что причиной гибели большого количества нейронов стало избыточное поступление ионов Са2+ в клетку в ходе следующих событий: ишемия > избыточное поступление ионов Са2+ в клетку > активация Са2+-зависимых протеаз и липаз > разрушение белков и липидов > гибель клетки. Назначение антагониста NMDA-рецепторов сразу же после первичного инсульта позволяет снизить тяжесть ишемического поражения мозга.
Метаботропные глутаматные рецепторы Выделяют более 100 различных метаботропных глутаматных рецепторов. Все метаботропные рецепторы — это внутренние мембранные белки, большинство которых располагается на постсинаптических мембранах и оказывает возбуждающее действие. Некоторые метаботропные рецепторы локализуются на пресинаптической мембране и являются тормозными ауторецепторами.
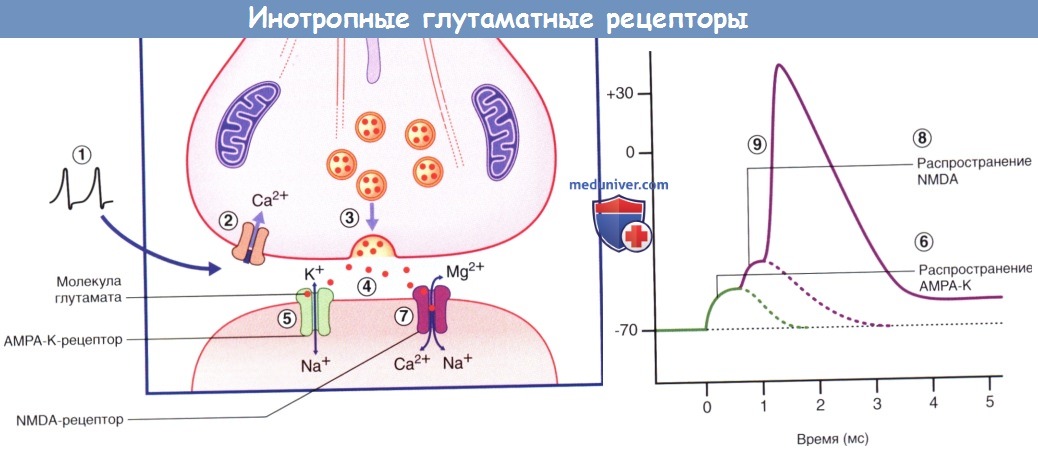
Ионотропные глутаматные рецепторы.
(1) При возникновении потенциала действия в области нервного окончания происходит (2) открытие кальциевых каналов (Ca2+).
(3) Под влиянием ионов Ca2+ синаптические пузырьки приближаются к плазматической мембране.
(4) Молекулы глутамата высвобождаются в синаптическую щель путем экзоцитоза.
(5) Медиатор связывается с AMPA-K-рецепторами, что вызывает открытие ионных каналов и поступление большого количества ионов Na+ в клетку, а также выход небольшого количества ионов К+ из клетки в результате чего (6) возникает возбуждающий постсинаптический потенциал (ВПСП), вызывающий деполяризацию значением 20 мВ, что делает возможной (7) активацию NMDA-рецептора глутаматом за счет «выталкивания» из ионного канала рецептора иона Mq24 Через канал NMDA-рецептора проникают ионы Na+ и Са2+, что приводит к деполяризации клетки.
(8) ВПСП, генерированный NMDA-рецептором, достаточен для (9) усиления потенциалов действия с продолжительным периодом реполяризации за счет повышения внутриклеточной концентрации ионов Са2+.
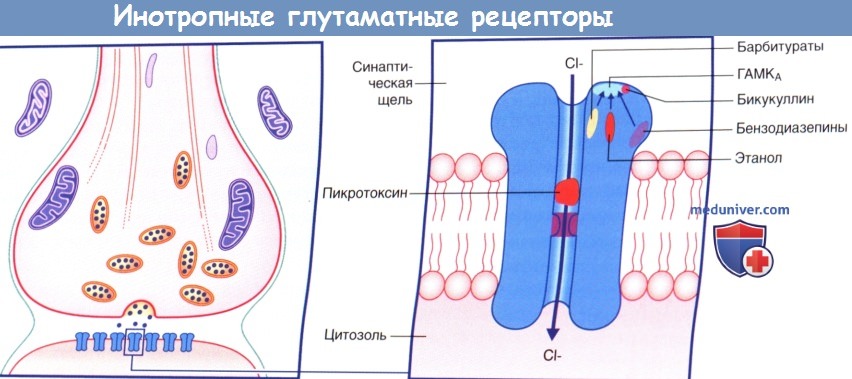
Лекарственные средства и ионотропный ГАМКA-рецептор. Зеленым цветом обозначено действие агонистов, красным цветом—действие антагониста.
Барбитураты, бензодиазепины и этанол вызывают гиперполяризацию клетки за счет воздействия на рецептор.
Бикукуллин—антагонист рецептора. Пикротоксин оказывает прямое действие, закрывая отверстие ионного канала.
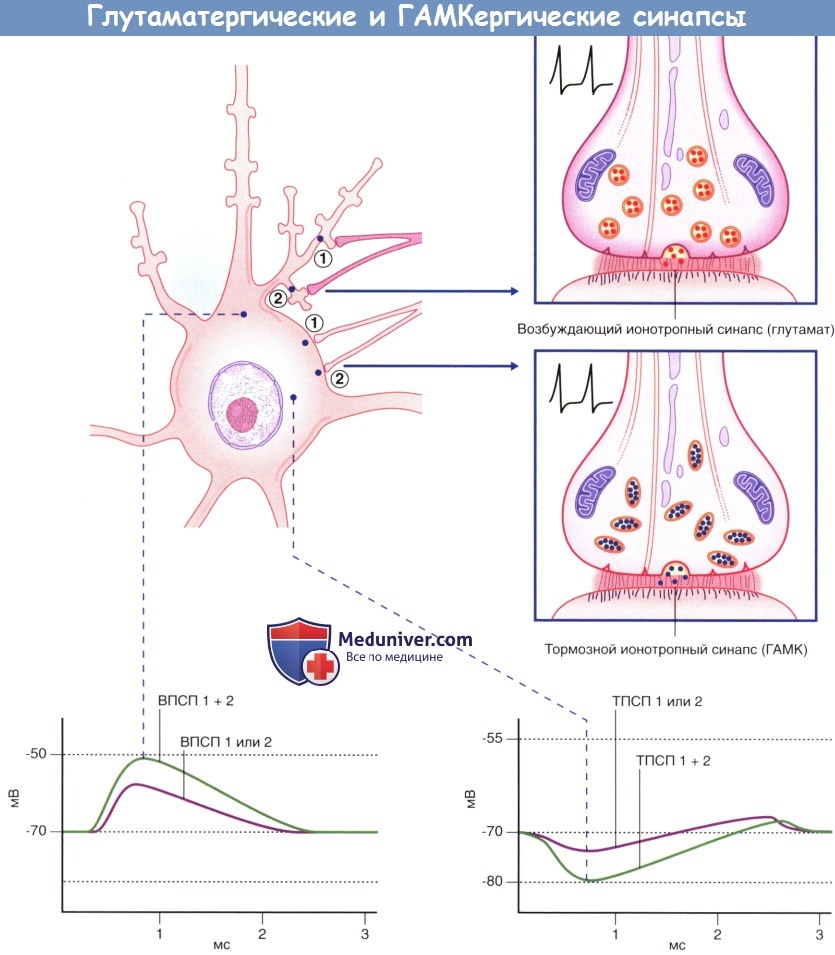
Глутаматергический и ГАМКергический синапсы мультиполярного нейрона с шипиковыми дендритами.
Продемонстрирована пространственная суммация возбуждений для каждой пары синапсов.
б) ГАМК. ГАМК-рецепторы могут быть как ионотропными, так и метаботропными.
1. Ионотропные ГАМК-рецепторы. Рецепторы, называемые ГАМКA, расположены в большом количестве в области лимбической доли головного мозга. Каждый рецептор связан с хлорным каналом. При активации ГАМКA-рецепторов хлорные каналы открываются, и ионы Cl- по градиенту концентрации поступают из синаптической щели в цитозоль. Причиной гиперполяризации, при которой достигаются значения -70 мВ и ниже, служит суммация последовательных ТПСП.
Действие седативных снотворных препаратов барбитуровой кислоты и бензодиазепина (например, диазепама) реализуется за счет активации ГАМКA-рецепторов. Аналогично действие этанола (потеря контроля социального поведения под влиянием действия этанола происходит вследствие растормаживания возбуждающих нейронов-мишеней, которые в обычном состоянии «сдерживаются» под действием ГАМКергических влияний). Механизм действия некоторых летучих анестетиков также заключается в связывании рецепторов, за счет чего ионные каналы остаются открытыми более долгое время.
Основной антагонист, занимающий активный центр рецептора, — конвульсант бикукуллин. Другой конвульсант — пикротоксин — связывается с субъединицами белка, в активном состоянии закрывающими ионный канал.
2. Метаботропные ГАМК-рецепторы. Метаботропные рецепторы, получившие название ГАМКВ, равномерно распределены во всех структурах мозга, их также обнаруживают в периферических вегетативных нервных сплетениях. Несмотря на то, что большое количество G-белков этих рецепторов выполняет роль вторичных посредников, значительная часть G-белков оказывает влияние на особый вид постсинаптических калиевых каналов — GIRK-каналы (G-белок-связанные калиевые каналы внутреннего выпрямления). При присоединении медиатора происходит отделение β-субъединицы, которая «выталкивает» ионы К+ через GIRK-канал, что приводит к формированию ТПСП.
Ответная реакция этого вида рецепторов нейрона-мишени медленнее и слабее по сравнению с ионофорезом ГАМКA, и для их активации необходима стимуляция большей частоты. В связи с этим считают, что ГАМКA-рецепторы расположены не во внешнем слое синаптической щели, а внесинаптически. Это предположение может быть подтверждено наличием еще одного вида расположенных внесинаптически G-направленных каналов. Эти кальциевые каналы также потенциалозависимые и принимают участие в обеспечении клетки количеством ионов Са2+, необходимым для перемещения синаптических пузырьков через пресинаптическую мембрану. При активации G-Са2+-лигандного участка связывания происходит закрытие кальциевых каналов, что приводит к снижению влияния потенциала действия, а также к торможению исходного нейрона (источника возбуждения) и других прилежащих глутаматергических нейронов.
В некоторых случаях для лечения заболеваний, связанных с чрезмерным рефлекторным тонусом мышц (мышечная спастичность), применяют инъекции миорелаксанта баклофена (агониста ГАМКВ) в окружающее спинной мозг субарахноидальное пространство. Баклофен проникает в спинной мозг и ингибирует высвобождение глутамата из чувствительных нервных окончаний в основном за счет уменьшения поступления большого количества ионов Са2+ в клетку, возникающего под влиянием потенциалов действия чрезмерной частоты.
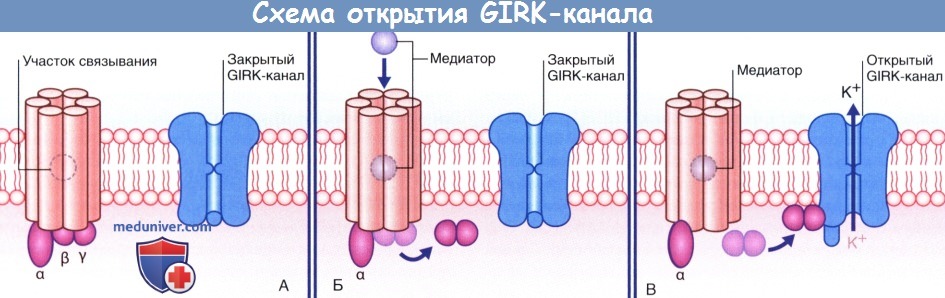
Схема открытия GIRK-канала, расположенного на постсинаптической мембране, G-белком.
(А) Состояние покоя. (Б) ГАМК активирует рецептор, и βγ-субъединица G-белка перемещается по направлению к GIRK-каналу.
(В) βγ-субъединица вызывает высвобождение ионов К+, что приводит к гиперполяризации мембраны.
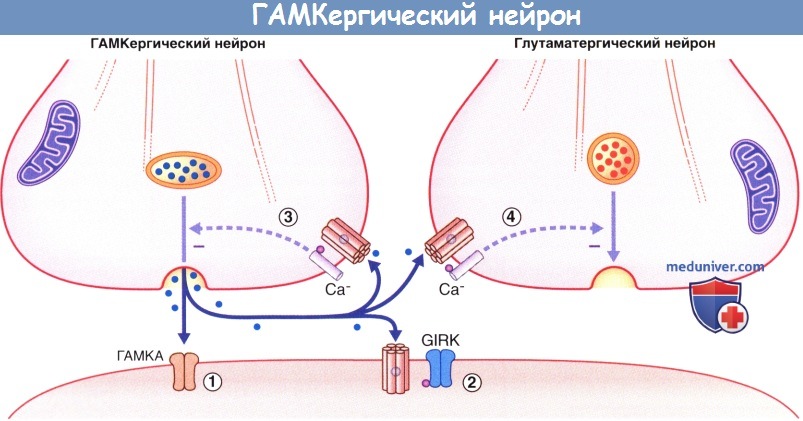
Высвобождение медиатора и дальнейшие процессы, происходящие в ГАМКергическом нейроне.
(1) Связываясь с ГАМКA-рецепторами, медиатор вызывает гиперполяризацию мембраны нейрона-мишени за счет открытия хлорных (Cl-) каналов.
(2) Аналогичное действие оказывают связывающиеся с GIRK рецепторы ГАМКВ за счет открытия G-белок-связанных калиевых каналов внутреннего выпрямления (GIRKS).
(3) При связывании ГАМКВ-ауторецепторов уменьшается высвобождение медиатора исходным нейроном за счет закрытия лиганд-G-белок-зависимых кальциевых (Са2+) каналов.
(4) Связывание ГАМКВ-рецепторов соседнего глутаматергического нейрона оказывает аналогичное влияние, опосредованное действием ионов Са2+.
3. Обратный захват глутамата и ГАМК. Обратный захват глутамата и ГАМК происходит двумя путями. В левой части каждого рисунка показано, что некоторые молекулы медиатора захватываются из синаптической щели транспортными белками мембраны и помещаются обратно в синаптические пузырьки. В правых частях рисунков изображен захват молекул медиаторов прилежащими астроцитами. Находясь в астроците, глутамат под действием глутаминсинтетазы превращается в глутамин. В процессе последующего транспорта к синаптическому уплотнению глутамат достраивается под действием глутаминазы и помещается в синаптический пузырек. ГАМК превращается в глутамат под действием ГАМК-трансаминазы. В процессе транспорта глутамат трансформируется в глутамин под действием глутаминсинтетазы.
Вернувшись в область синаптического уплотнения, глутамин под действием глутаминазы превращается в глутамат, из которого под действием глутаматдекарбоксилазы синтезируется ГАМК, молекулы которой помещаются в синаптические пузырьки.
Блокирование фермента глутаматдекарбоксилазы лежит в основе известного аутоиммунного заболевания — синдрома «скованного человека».
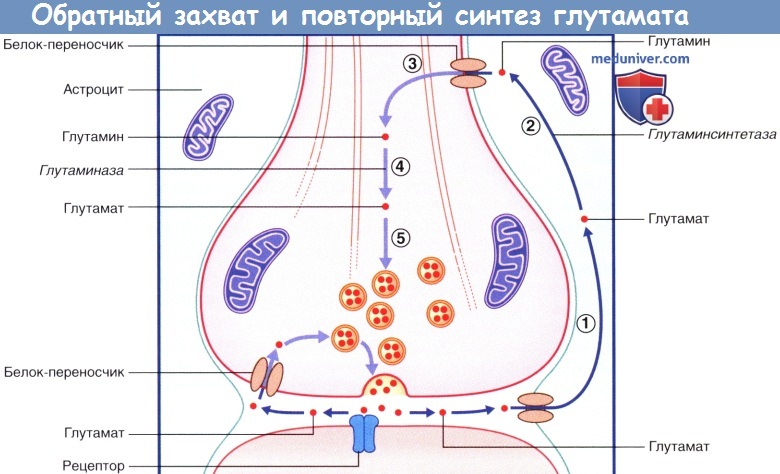
Схема обратного захвата и повторного синтеза глутамата.
В левой части рисунка происходит обратный захват молекулы глутамата в неизменном виде.
В правой части рисунка (1) глутамат захватывается астроцитами, затем (2) под действием глутаминсинтетазы превращается в глутамин.
(3) Глутамин поступает в нервное окончание, (4) где под действием глутаминазы превращается в глутамат, который (5) возвращается в синаптические пузырьки.
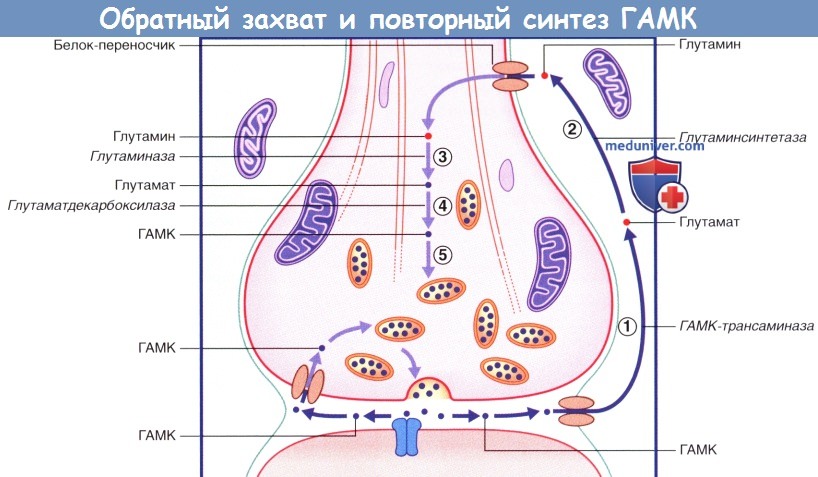
Схема обратного захвата и повторного синтеза ГАМК. В левой части рисунка происходит обратный захват молекулы ГАМК в неизменном виде.
В правой части рисунка ГАМК захватывается астроцитами, затем (1) под действием ГАМК-трансаминазы превращается в глутамат, который (2) под действием глутаминсинтетазы превращается в глутамин.
(3) Глутамин поступает в нервное окончание и под действием глутаминазы образует глутамат.
(4) Глутамат под действием глутаматдекарбоксилазы превращается в ГАМК, которая (5) возвращается в синаптические пузырьки.
г) Глицин. Глицин синтезируется из серина в процессе катаболизма глюкозы. Основная функция этого нейромедиатора — обеспечение отрицательной обратной связи двигательных нейронов ствола мозга и спинного мозга. При инактивации глицина (например, при отравлении стрихнином) возникают мучительные судороги.
Обратный захват. В области синаптического уплотнения при помощи аксональных белков-переносчиков осуществляется быстрый обратный захват глицина с последующим его помещением в синаптические пузырьки.
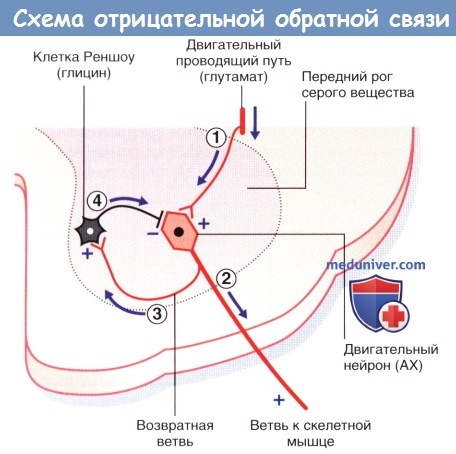
Схема отрицательной обратной связи: клетки Реншоу ингибируют избыточное возбуждение двигательных нейронов. АХ—ацетилхолин.
(1) Нейрон нисходящего двигательного проводящего пути оказывает возбуждающее действие на двигательный нейрон спинного мозга.
(2) Двигательный нейрон вызывает сокращение мускулатуры.
(3) Возвратная ветвь стимулирует клетку Реншоу.
(4) Клетка Реншоу оказывает ингибирующее влияние, достаточное для предупреждения чрезмерной активации двигательного нейрона.