• Кэпирующие белки функционируют как на оперенных так и на заостренных концах актиновых филаментов
• Кэпирующие белки и белки семейства гельзолин подавляют элонгацию оперенных концов; в свою очередь, эта их способность ингибируется фосфолипидами плазматической мембраны
• Тропомодулин представляет собой белок, который кэпирует заостренные конец актинового филамента
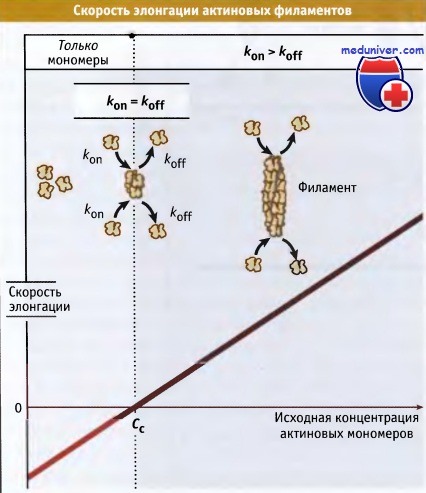
In vitro, при нерегулируемом росте, актиновые филаменты быстро и непрерывно растут до тех пор, пока концентрация свободного мономера не достигнет критического уровня. В клетке существуют специальные механизмы, которые контролируют количество свободных оперенных концов филаментов. Такой контроль необходим для предотвращения истощения пула мономерного актина и для регулировки размера специфических актиновых структур.
Поскольку короткие актиновые филаменты обладают большей жесткостью, чем длинные, регуляция их размеров контролирует механические свойства актиновой сети. Белки, которые связываются с концами актиновых филаментов и предотвращают дальнейшую полимеризацию мономеров, называются кэпирующими белками. Некоторые кэпирующие белки связывают оперенные концы, в то время как другие ассоциируют с заостренными концами. Белки, образующие кэп на оперенном конце, ограничивают длину филамента, поскольку препятствуют элонгации. Белки, кэпирующие заостренный конец, предотвращают деполимеризацию актина.
К белкам, образующим кэп на оперенном конце, относятся CapZ, EPS8 и представители семейства гельзолин. Хотя кэпирующие и гельзолин-белки различаются по структуре и механизму действия, они связываются с оперенными концами филаментов и делают невозможным дальнейшее добавление мономеров. Эти белки обладают высоким сродством к оперенным концам филаментов и препятствуют добавлению актиновых субъединиц даже при высокой концентрации мономеров.
Свойства белков и копирующих белков гельзолина регулируются фосфолипидами плазматической мембраны. Фосфолипид фосфатидилинозитол 4,5-дифосфат (PIP2) разрушает связь между обоими типами копирующих белков и оперенными концами актиновых филаментов. Поэтому они обеспечивают декэпирование филаментов или предотвращают их копирование на плазматической мембране. Фосфолипид PIP2 расположен во внутреннем слое плазматической мембраны и участвует во внутриклеточной системе передачи сигналов.
Уровень PIP2 меняется в зависимости от сигнала, поступающего с поверхностных рецепторов клетки, сопряженных с одним из G-белков. Путем регуляции копирования на клеточной мембране можно контролировать элонгацию филаментов. Этот процесс является ключевым в миграции клеток и протрузии мембран. Уровень активности копирующих белков может влиять на тип протрузии (ламеллоподии или филоподии).
Тропомодулины представляют собой семейство широко распространенных копирующих белков, которые в присутствии регуляторного актинового белка тропо-миозина обладают высоким сродством к заостренным концам актиновых филаментов. Тропомодулины контролируют длину актиновых филаментов волокон поперечно-полосатых мышц. Они также регулируют длину актиновых филаментов и динамику их сборки в эритроцитах и эпителиальных клетках
По данным экспериментов in vitro, скорость элонгации линейно зависит от концентрации актиновых мономеров.
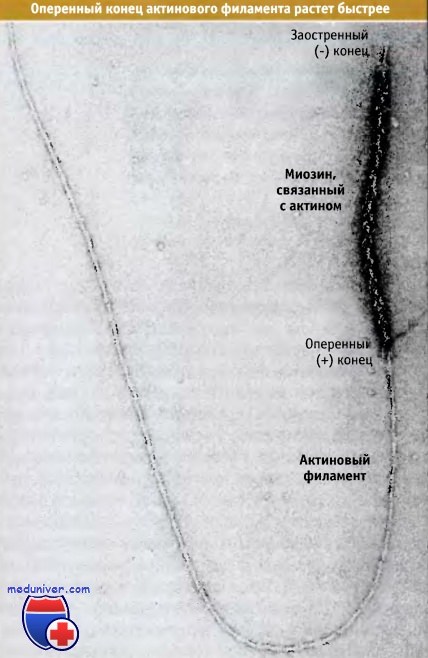
Результаты эксперимента, демонстрирующего, что полимеризация актина, главным образом, происходит на оперенном конце филамента.
Для нуклеации процесса элонгации использовали филамент, связанный с актином.
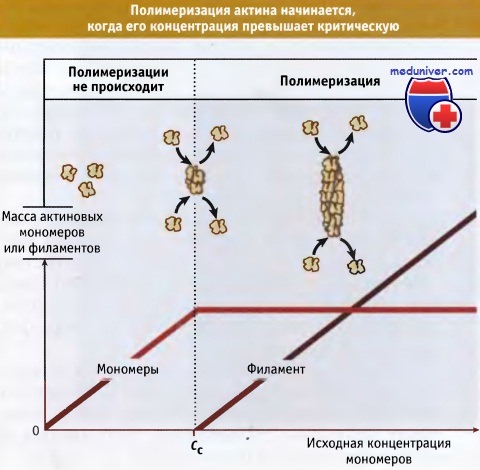
По данным исследований in vitro, сборка акти-новых филаментов начинается,
когда исходная концентрация актиновых мономеров превышает критическую концентрацию, Сс.