• Белки нуклеации позволяют клетке контролировать время и место образования филаментов de novo
• In vivo нуклеация филаментов происходит с участием комплекса Arp2/3 и форминов
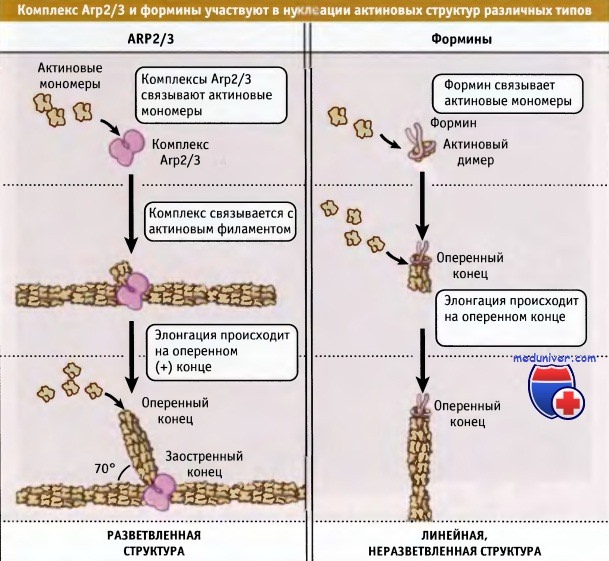
• При нуклеации с участием Arp2/3 образуется разветвленная сеть филаментов, а форминовые белки инициируют неразветвленные филаменты
• Arp2/3 активируется на клеточной мембране при действии белков Scar, WASP и WAVE
Нуклеация является наиболее медленным этапом образования нового актинового филамента in vitro и важнейшим средством контроля за регуляцией полимеризации клеточного актина. Для быстрой пространственно регулируемой полимеризации нового актинового филамента этап нуклеации необходимо ускорить. Белки, которые облегчают образование филаментов de novo, называются белками нуклеации.
Комплекс Arp2/3 и формины хорошо охарактеризованы и представляют собой два типа белков нуклеации актиновых филаментов, играющих важную роль в регуляции подвижности клеток. Эти белки нуклеируют филаменты по разным механизмам и способствуют образованию в клетке различных типов сети филаментов.
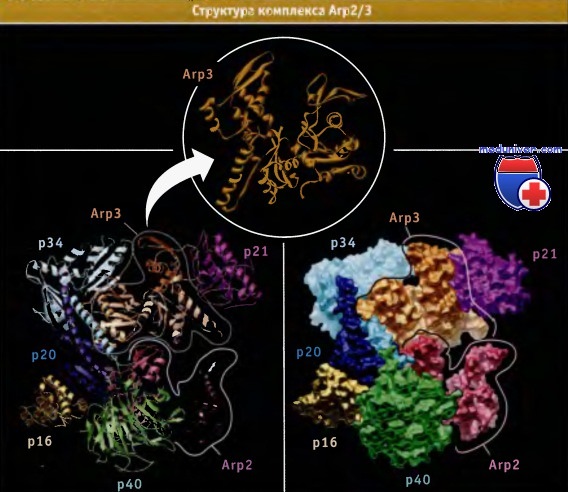
По данным рентгеноструктурного анализа, Arp2/3 представляет собой макромолекулярный белковый комплекс, состоящий из Arp2, ArpЗ и пяти дополнительных белков. Arp2 и Arp3 напоминают актин. Хотя по структуре эти белки близки к актину, они не могут полимеризоваться с образованием филаментов. Предполагается, что Arp2 и Arp3 связываются в комплекс таким образом, что он становится похож на стабильный димер актина с открытым оперенным концом.
Это помогает стабильному нуклеусу образоваться в момент, когда актиновый мономер связывается с комплексом. По мере элонгации нового филамента на оперенном конце, комплекс Arp2/3 остается связанным с заостренным концом.
Способность комплекса Arp2/3 к нуклеации усиливается при его взаимодействии с регуляторными белками и с боковыми поверхностями предсуществующих актиновых филаментов. Нуклеация может происходить на плазматической мембране. Новообразованные «дочерние» филаменты растут в направлении оперенного конца, подобно веткам располагаясь под углом 70° по бокам предсуществующих «материнских» филаментов.
Как следует из экспериментов in vitro, способность комплекса Arp2/3 к связыванию с боковыми поверхностями филаментов приводит к образованию разветвленной сети актиновых фибрилл, напоминающую ту, которая наблюдается на быстрорастущих краях подвижных клеток. Комплекс Arp2/3 связывается более прочно с филаментами тех субъединиц, которые содержат АТФ или АДФ-Фн («молодые» филаменты), чем с субъединицами, содержащими АДФ («старые» филаменты). Поэтому Arp2/3 преимущественно связывается с молодыми филаментами и образует на них ветвящиеся структуры. После высвобождения Фг и старения филамента Arp2/3 отщепляется. Это приводит к диссоциации боковых ветвей и к разборке актиновой сети.
Посредством нескольких белков, система передачи сигнала активирует комплекс Arp2/3 на мембранах. Активация Arp2/3 имеет важное значение для роли, которую играет актин в протрузии мембран. Эти белки активируются малыми G-белками и включают Scar (супрессор рецептора цАМФ), WASP (Wiskott-Aldrich syndrome protein) и WAVE (WASP-^crprolin homologs). Вместе с существующими актиновыми филаментами они обеспечивают активацию комплекса Arp2/3.
Формины относятся к семейству структурно близких белков. Своим названием они обязаны мышиному гену, limb deformity. Эти белки содержат два уникальных гомологичных домена, которые называются формин гомологичный-1 и -2 (FH1 и FH2). Домен FH1 связывает профилин, a FH2 участвует в процессе нуклеации полимеризации актина. Формины характеризуются рядом примечательных особенностей: в ходе элонгации оперенного конца филамента они остаются связанными с ним; защищают его от копирующих белков при элонгации и увеличивают ее скорость за счет прямой ассоциации с профилином.
Механизм нуклеации и элонгации с участием форминов отличается от механизма действия комплекса Arp2/3, который при элонгации остается на заостренном конце филамента. Поскольку, в отличие от разветвленной сети актина, создаваемой комплексом Arp2/3, формины нуклеируют образование актиновых филаментов, не связываясь с боковыми поверхностями уже существующих, они регулируют сборку неразветвленных филаментов.
Кристаллическая структура белкового комплекса Arp2/3. Представлена третичная структура каждой субъединицы (слева) или ее объемная модель (справа).
Белки Arp2 и Arp3 по структуре близки к актину. Структура Arp3 построена по данным из Protein Data Bank file 1KBK.
Комплекс Arp2/3 обеспечивает нуклеацию разветвленных актиновых структур,
в то время как формины участвуют в нуклеации неразветвленных, линейных актиновых образований.