• Во вногих эукариотических клетках присутствуют два белка, которые связываются с мономерным актином. Это тимозин b4 и профилин
• В клетках Metazoa тимозин b4 изолирует актиновые мономеры и поддерживает цитозольный пул АТФ-актина, который может быть использован для быстрой элонгации филаментов
• Комплексы профилина с актиновыми мономерами способствуют элонгации только оперенных концов филаментов
В клетках поддерживается пул мономерного актина, который может быть использован для быстрой элонгации, когда внешний сигнал определяет время и место запуска процесса роста филаментов. Скорость элонгации актинового филамента зависит от концентрации актиновых мономеров, доступных для полимеризации. Для того чтобы осуществить быструю элонгацию в соответствующей клетке шкале времени, концентрация актиновых мономеров, доступных для полимеризации, должна быть гораздо выше критической. Белки, которые связываются с актиновыми мономерами, способствуют регуляции скорости роста филаментов.
В клетках Metazoa присутствуют два наиболее распространенных белка, которые связываются с мономерами актина. Это тимозин b4 и профилин. Тимозин b4 обнаружен только у высших эукариот и, главным образом, находится в высокоподвижных и способных к фагоцитозу клетках. Профилин найден в большинстве эукариот, включая клетки растений, животных и дрожжей.
Концентрация этих белков сопоставима с клеточной концентрацией актина. Хотя оба белка связывают актиновые мономеры, тимозин b4 и профилин по-разному регулируют полимеризацию актина, и оба играют критическую роль в физиологических процессах.
Тимозин b4 представляет собой небольшой пептид (Мr < 5кДа). Он принадлежит к семейству белков, связывающихся с актиновым мономером, и образующих с ним комплекс в соотношении 1:1. Эти белки обладают консервативной первичной структурой. Тимозин b4 ингибирует спонтанную полимеризацию актиновых мономеров и предотвращает добавление мономеров к существующим филаментам.
Хотя белок представляет собой небольшой пептид, считается, что он связывается с протяженным участком поверхности мономера актина, стерически блокируя много областей, необходимых для образования важных контактов, стабилизирующих филаменты. В клетках, обладающих повышенной подвижностью, таких как нейтрофилы, концентрация тимозина b4 может достигать нескольких сотен микромоль. Это позволяет таким клеткам поддерживать большой цитоплазматический пул мономера актина. Тимозин b4 связывается с более высоким сродством с АТФ-актиновыми (Кd ~2 мкМ), чем с АДФ-актиновыми мономерами (Kd ~50 мкМ). Поэтому пул неполимерного актина в клетках состоит, главным образом, из АТФ-актина.
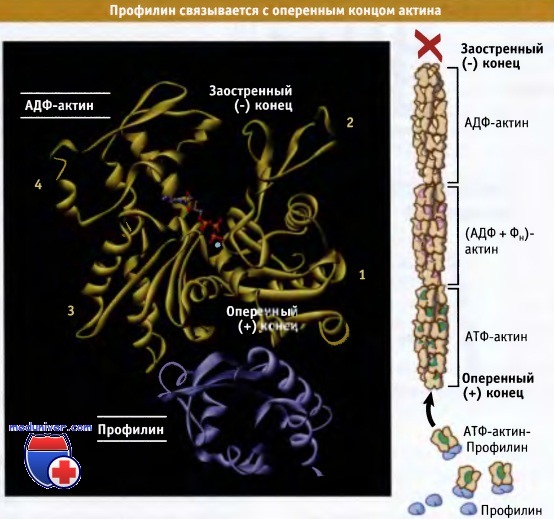
Профилин также является небольшим (Мr ~ 15 кДа) белком, способным связываться с мономерным актином, образуя комплексы в соотношении 1:1. Однако, в отличие от тимозина b4, эти комплексы могут связываться с оперенными концами актиновых филаментов. При связывании комплекса с оперенным концом профилин диссоциирует и в дальнейшем может связать другой мономер актина. Рисунок ниже показывает, что поскольку профилин связывает мономеры актина по субдоменам 1 и 3, в его присутствии блокируется связывание мономера с заостренным концом филамента. Таким образом, комплекс профилин-актин способствует элонгации исключительно на оперенном конце филамента.
Еще одно ключевое свойство профилина — это его способность обменивать нуклеотид, связанный с мономерным актином. Поскольку, внутриклеточная концентрация АТФ гораздо выше, чем АДФ, присутствие профилина позволяет мономеру актина быстро вступать в равновесие с цитоплазматическим АТФ и превращать АДФ-актиновые мономеры в АТФ-актиновые. Это способствует поддержанию большого пула АТФ-актиновых мономеров, необходимых для быстрого роста филаментов.
Профилин связывается с оперенным концом актиновых мономеров
и обеспечивает регуляцию полимеризации актина за счет включения мономера только со стороны оперенного конца.