• Рост и организация структур клеточных отростков регулируются взаимодействием между актином и белками, взаимодействующими с его мономерами и филаментами
• При добавлении актиновых мономеров к оперенным концам актиновых филаментов, расположенных на плазматической мембране, она сдвигается кнаружи (в направлении роста)
Для перемещения клетки по поверхности необходимо формирование и рост клеточных отростков богатых актином, фиксация этих структур на поверхности, и сокращение тела клетки. В настоящей статье мы опишем, каким образом за счет полимеризации актина и перестройки цитоскелета происходит выпячивание переднего участка ламеллоподии мигрирующей клетки, представленное на рисунке ниже.
Механизм формирования и роста ламеллоподий хорошо изучен. Для образования ламеллоподий необходима нуклеация актиновых филаментов с участием комплекса Arp2/3. При последующем добавлении актиновых мономеров к оперенному концу актинового филамента с краю клетки плазматическая мембрана выпячивается вперед. Отметим, что подвижность клеток можно наблюдать в системе in vitro, содержащей очищенный актин, активированный Arp2/3, кофилин и копирующий белок.
Во многих клетках, по мере того как они перемещаются вперед, поддерживается относительно постоянная величина ламеллоподий. Это объясняется тем, что скорость добавления мономеров на переднем участке ламеллоподий такая же, как скорость разборки филамен-тов на их заднем участке. Рост отростков и их геометрия регулируются взаимодействием между актином, белками нуклеации и кэпирования, а также белками, регулирующими их функции и разрезающими филаменты.
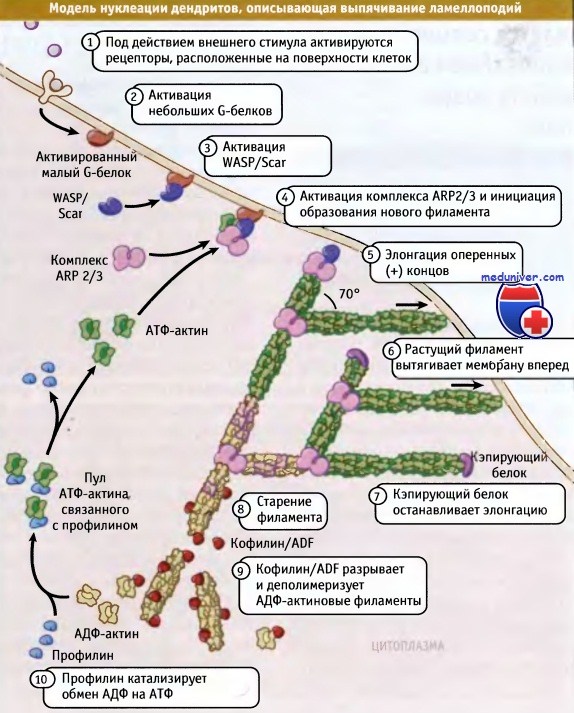
Цепочка событий, приводящих к перестройке актинового цитоскелета, начинается с внешнего сигнала, запускающего систему его реализации в клетке, которая приводит к активации белков WASP/Scar.
Эти белки связывают и активируют комплексы Arp2/3, которые нуклеируют новые филаменты, ответвляющиеся от материнских. Геометрия активации обеспечивает ориентацию оперенных концов новых филаментов в направлении плазматической мембраны.
Филаменты быстро удлиняются и сдвигают мембрану вперед, до тех пор пока оперенные концы не копируются соответствующими белками. Комплекс Arp2/3 и новообразованные короткие копированные актиновые филаменты образуют сетчатую структуру, которая с механической точки зрения хорошо приспособлена к постоянной протрузии мембраны по поверхности.
Необходимые для элонгации мономеры поступают из большого пула растворимых комплексов АТФ-актина и тимозина b4. АТФ-актиновые мономеры находятся в состоянии равновесия с тимозином b4 и профилином. Актин в профилин-актиновом комплексе связан со свободными оперенными концами актиновых филаментов. При связывании профилин-актинового комплекса с оперенным концом происходит диссоциация профилина.
Это создает возможность добавления к филаменту следующего мономера. АТФ, связанный с полимеризованными субъединицами актина, гидролизуется и происходит медленное высвобождение фосфата, которое приводит к образованию АДФ-актина. За счет высвобождения фосфата увеличивается скорость диссоциации комплекса Arp2/3; при этом кофилин/ADF начинает связывать АДФ-актин, а также разрезать и деполимеризовать актиновые филаменты.
Затем профилин начинает катализировать обмен АДФ на АТФ в актиновых мономерах, которые высвобождаются из филаментов. После этого АТФ-актиновые мономеры связывают тимозин b4 и пополняют пул мономерного актина.
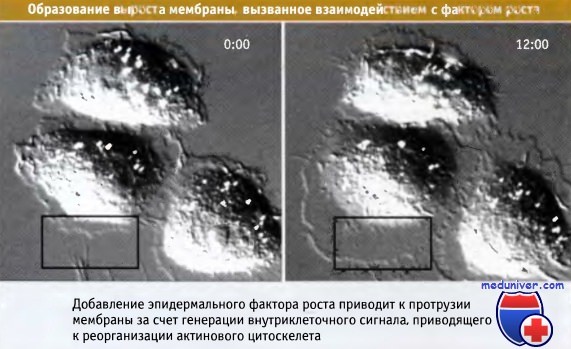
Эпидермальный фактор роста (EGF) стимулирует миграцию и деление клеток, экспрессирующих рецепторы EGF.
На этих видеокадрах видны эпителиальные клетки, у которых после добавления EGF наблюдается образование выроста мембраны
— протрузии мембраны, связанное с полимеризацией актина. Время в мин:с.
В ответ на внешний сигнал клетка может начать мигрировать в определенном направлении.
Модель нуклеации дендритов описывает, каким образом Arp2/3 энуклеирует сборку сети разветвленных актиновых филаментов,
и как в процессе этой сборки край клетки выдвигается вперед.