Механизм гидролиза АТФ актином после полимеризации
• Гидролиз АТФ под действием субъединиц актинового филамента носит необратимый характер. Поэтому полимеризация актина является неравновесным процессом
• Значение критической концентрации для сборки актина зависит от того, какой нуклеотид, АТФ или АДФ, связан с мономером
• Критическая концентрация для АТФ-актина ниже, чем для АДФ-актина
• В присутствии АТФ для двух концов актинового филамента существуют две различные критические концентрации
Как описано в предыдущих статьях на сайте, процесс спонтанной сборки актина in vitro может быть достаточно точно описан с помощью стандартных представлений, основанных на законе действующих масс и кинетике обратимых химических реакций. Однако актиновые субъединицы обладают ферментативной активностью и после включения в филамент гидролизуют связанный АТФ. Это вносит дополнительную сложность в описание процесса сборки актина. Мономерный актин не гидролизует АТФ, однако после полимеризации приобретает такую способность. При этом образуется АДФ и неорганический фосфат (Фн). Процесс гидролиза АТФ является практически необратимым. После гидролиза АДФ и Фн остаются связанными с актином нековалентными связями. Фосфат медленно высвобождается из филамента, однако АДФ остается прочно с ним связанным и не отщепляется в течение всего времени, пока субъединица находится в составе филамента.
Для полимеризации не требуется гидролиз АТФ. Фактически при полимеризации актина между включением субъединиц в филамент и гидролизом АТФ существует некоторый лаг-период. Более того, сам АТФ не нужен для полимеризации: АДФ-актиновые мономеры способны сами собираться в филаменты. Однако гидролиз АТФ необходим для регуляции функций актина в клетке.
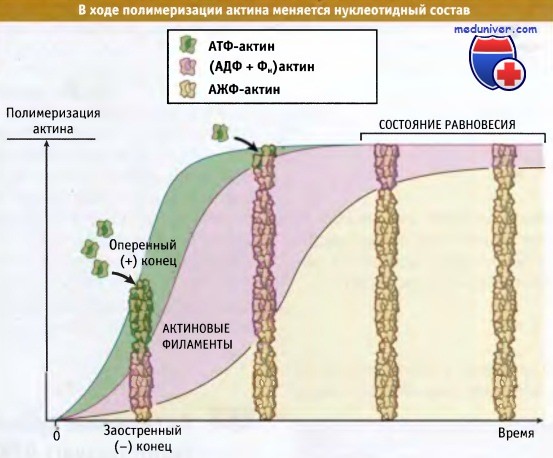
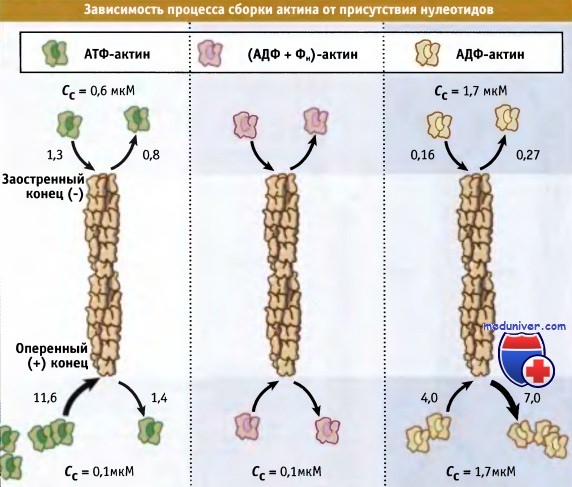
Как следует из рисунка ниже, критическая концентрация для процесса сборки актина зависит от природы связанного нуклеотида (АТФ, АДФ-Фн, или АДФ). Значение критической концентрации определяется концентрацией мономерного актина, необходимой для образования филаментов и рассчитывается из значений koff и kon, которые определяются в системе in vitro. На оперенном конце филамента критическая концентрация АТФ-актина (0,1 мкМ) оказывается ниже, чем АДФ-актина (1,7 мкМ). Это означает, что очищенные АТФ-актиновые мономеры будут полимеризоваться в филаменты до тех пор, пока их концентрация не достигнет 0,1 мкМ.
Аналогичным образом, АДФ-актин будет полимеризоваться до тех пор, пока его концентрация не достигнет 1,9 мкМ. Критическая концентрация АДФ-ФН актина на оперенном конце (0,1 мкМ) сопоставима с концентрацией АТФ-актина. Поскольку Сс = kd, эти значения указывают, что при высвобождении Фн происходят изменения в прочности связывания терминальной субъединицы. Иными словами, энергия гидролиза АТФ запасается в филаменте до высвобождения Фн, после чего терминальные субъединицы (уже содержащие АДФ) оказываются связанными с филаментом более слабой связью.
Для актиновых филаментов, содержащих АДФ-актин, критическая концентрация АДФ-актиновых мономеров на обоих концах филамента примерно одна и та же (2 мкМ). Это связано с тем, что субъединицы в филаменте и стабилизирующие контакты между ними одинаковы. Однако в присутствии АТФ критическая концентрация на оперенном конце (0,1 мкМ) оказывается ниже, чем на заостренном (0,7 мкМ). Это объясняется тем, что актиновые субъединицы на оперенном конце содержат АТФ (или АДФ-Фн), а на заостренном конце связанный АДФ. Такая химическая поляризация филамента обусловлена быстрым добавлением АТФ-мономеров к оперенному концу, что приводит к образованию «кэпа из АТФ». На заостренном конце добавление мономера происходит медленнее, так что гидролиз АТФ и высвобождение фосфата происходят быстрее, чем добавление АТФ-актиновых мономеров (если полимеризация не происходит очень быстро).
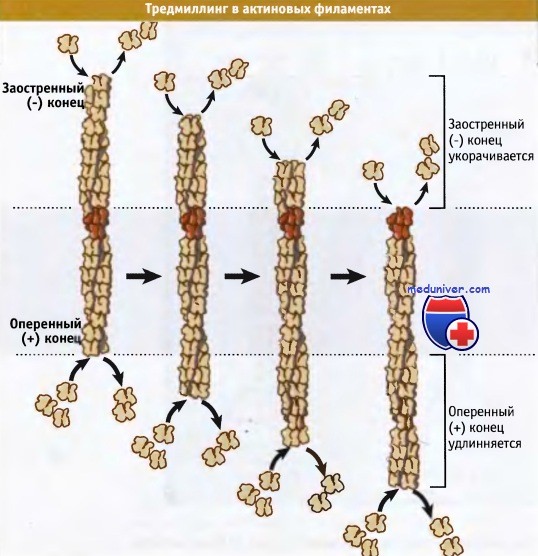
Когда пул актиновых мономеров представлен АТФ-актином (что характерно для клеток), критическая концентрация для всего филамента (т. е. равновесная концентрация актинового мономера) лежит между критическими концентрациями для двух концов: выше, чем для оперенного, и ниже, чем для заостренного конца. Следовательно, в состоянии равновесия актиновые мономеры (в виде АТФ-актина) связаны с оперенным концом и после гидролиза АТФ и высвобождения Фн отщепляются от заостренного конца в виде АДФ-актина. Процесс полимеризации-деполимеризации обусловливают постоянный поток субъединиц через филаменты, что не сопровождается изменениями общей концентрации полимера или мономера. Этот поток называется тредмиллингом (круговорот мономеров) и представлен на рисунке ниже. В клетках не происходит нерегулируемый тредмиллинг; скорее всего, процесс контролируется белками, связывающимися с актином.
В экспериментальных условиях, когда филаменты собираются из актиновых мономеров, содержащих АДФ, тредмиллинга не происходит, поскольку критическая концентрация на обоих концах филамента одинакова. Таким образом, актиновые субъединицы, отщепляющиеся от заостренного конца, должны обменивать связанный АДФ на АТФ, чтобы присоединиться к оперенному концу. Энергия гидролиза АТФ используется не для самого процесса полимеризации, а для обеспечения сборки и разборки актина и направленного движения мономеров по филаментам.
Тредмиллинг происходит только в том случае, когда концентрация актиновых мономеров находится между значениями критических концентраций на концах филамента. Если концентрация мономера на оперенном и заостренном концах оказывается выше критической, то происходит элонгация филаментов с обоих концов. Филаменты диссоциируют с обоих концов, когда концентрация мономера становится ниже критической. При концентрации мономера, занимающей промежуточное положение между критическими концентрациями на концах, мономеры добавляются к оперенным концам и отщепляются от заостренных. Таким образом, становится понятным, что если в клетке осуществляется контроль над концентрацией актиновых мономеров и критической концентрацией на обоих концах, то можно регулировать полноту, скорость и локализацию процессов полимеризации и диссоциации. Этот вопрос подробно обсуждается в следующем разделе.
Механизм полимеризации актиновых филаментов и микротрубочек имеет ряд общих черт. Прежде всего в результате обоих процессов собираются полярные структуры, и процесс элонгации сопряжен с гидролизом нуклеотидов. Для микротрубочек характерна динамическая нестабильность, при которой происходит быстрая полимеризация и диссоциация филаментов. Однако для актиновых филаментов нехарактерно существование фазы (фаза катастрофы), при которой происходит быстрая разборка филаментов. Это различие между актиновыми филаментами и микротрубочками обусловлено тем, что по сравнению с тубулином мономерный актин отщепляется от филамента в 100 раз медленнее.
Поскольку в начале фазы элонгации АТФ-актиновые мономеры связаны с оперенным концом, актиновые субъединицы содержат АТФ или, после гидролиза, АДФ и Фн.
По мере прохождения элонгации и истощения пула свободного мономера, процесс замедляется сильнее, чем гидролиз АТФ.
Таким образом, в состоянии равновесия актиновые филаменты содержат АДФ-актин, за исключением тех, на оперенном конце которых находится АДФ и Фн.
In vitro в состоянии равновесия критическая концентрация (Сс) для АТФ-актина или АДФ + Фн-актина на оперенном конце филамента оказывается ниже,
чем на заостренном конце, в то время как значения Сс для АДФ-актина на обоих концах филамента остаются одинаковыми.
Также представлены значения констант скорости для процессов ассоциации и диссоциации мономеров актина, kon и koff соответственно, которые используются для расчета Сс.
В состоянии равновесия в актиновых филаментах происходит тредмиллинг, при котором АТФ-актиновые мономеры быстрее присоединяются к оперенному концу,
а АДФ-актиновые мономеры быстрее диссоциируют от заостренного конца. При этом длина филамента не изменяется.