Связывание SRP с рецептором при закреплении белков на мембране ЭПР
• Причаливание (докинг) SRP к ее рецептору локализует рибосому и новообразующуюся цепь белка поблизости от транслокона
• Для причаливания необходимо связывание ГТФ и наличие гидролитической активности у SRP и ее рецептора
Узнавание новообразующейся цепи секреторного или мембранного белка при помощи SRP завершает только первую половину процесса адресования. Связавшись с SRP, белковая цепь должна позиционироваться на мембране ЭПР и транспортироваться к транслокационному каналу. В качестве промежуточного участника этого процесса выступает белковый комплекс, известный под названием SRP-рецептор (SR), который локализован на мембране ЭПР со стороны цитозоля.
SR представляет собой димер, состоящий из двух близких по строению субъединиц. Субъединица «альфа» ориентирована в сторону цитозоля (SR а), она взаимодействует с SRP, а «бета» (SR b) встроена в мембрану. Она взаимодействует с SR а и фиксирует ее на мембране ЭПР.
Подобно SRP54, каждая из субъединиц, SRa и SRb, содержит домен, ответственный за связывание и гидролиз ГТФ, и три этих белка образуют подсемейство ГТФ-аз. Ферментативная активность этих белков в отношении ГТФ играет критическую роль в адресовании белка на ЭПР и в его переносе в канал транслокации, а также в рециклировании SRP после прохождения адресования.
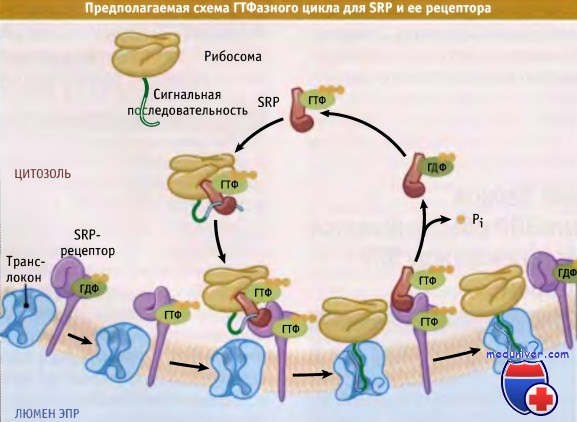
Как иллюстрирует рисунок ниже, для прохождения адресования необходимо, чтобы SRP и SR осуществляли скоординированное связывание и гидролиз ГТФ. Пока не выяснено, связывают ли SRP и SR а ГТФ до их ассоциации друг с другом или их взаимодействие приводит к связыванию ГТФ. В любом случае связанная с сигнальной последовательностью SRP и рибосома объединяются с SR. SR связывается с транслоконом, и объединение ГТФ с SRb, вероятно, облегчается субъединицей канала транслокации.
Более того, происходит связывание транслокона с комплексом «рибосома-растущая цепь». Таким образом, возникает продукт скоординированной ассоциации нескольких компонентов. В этом продукте SRP и обе субъединицы SR содержат связанный ГТФ. По-видимому, конформационные изменения, приводящие к гидролизу ГТФ под действием SRP и SR, возникают лишь в случае успешного образования комплекса, и как SRP, так и SR а способны взаимно активировать этот процесс, в результате чего их структура существенно меняется.
Рибосома также может ускорять гидролиз ГТФ в SRP и SR а. Изменения конформации приводят к высвобождению комплекса «рибосома-растущая цепь» из SRP и SR. В то же время за счет взаимодействия между рибосомой, каналом и белковой цепью последняя удерживается на месте и готова к транслокации.
Многократные взаимодействия белков и гидролиз нуклеотидов при адресовании, вероятно, обеспечивают необходимую скорость и степень точности этого процесса. Если при синтезе белка цитозоля рибосома располагается на мембране ЭПР поблизости от транслокона, то отсутствие ассоциации между SRP-SR свидетельствует о том, что белок находится здесь временно и не будет транслоцироваться. Аналогичным образом, если рибосома позиционирована на комплексе SR, который не связан с каналом, то взаимодействие не будет усилено из-за отсутствия связи рибосома-транслокон и транслокон-SR, поэтому гидролиз ГТФ происходить не будет. Таким образом, в отсутствие транслокона белковая цепь не будет освобождаться.
При высвобождении из SRP рибосома занимает транслокон. Это осуществляется при ее непосредственном взаимодействии с белками канала. При этом рибосома позиционируется непосредственно над входом в канал со стороны цитозоля, и новообразующийся полипептид транспортируется между рибосомой и каналом. Взаимодействие между ними сначала слабое, но может усиливаться, и существует на протяжении всего процесса транслокации.
Остается неясным один аспект адресования и транслокации. Каким образом присутствие нескольких рибосом на одной молекуле иРНК (полисома) влияет на адресование и сборку транслоконов. Если белок синтезировался с участием первой рибосомы на иРНК, позиционированной на ЭПР, то для остальных рибосом SRP должны быть не нужны, поскольку они уже были позиционированы около мембраны ЭПР.
Исследования, проведенные методом резонансного переноса энергии флуоресценции (FRET), который позволяет измерить расстояние между двумя молекулами, показали, что, несмотря на незначительные структурные различия между свободным и занятым транслоконом, канал остается собранным и готовым к транслокации, даже если не занят. Таким образом, взаимодействие SRP-SR может оказаться необходимым только для начального этапа адресования. После этого остальные рибосомы довольно быстро ассоциируют с транслоконами, и происходит узнавание сигнальной последовательности, что является как бы сверкой, которая обеспечивает уверенность в том, что каждый полипептид, подлежащий транслокации, действительно представляет собой секреторный или трансмембранный белок.
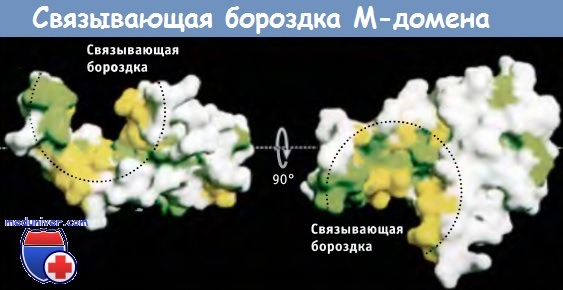
Строение М-домена бактериального гомолога SRP54.
Остатки гидрофобных аминокислот обозначены зеленым и желтым цветом.
Эти остатки образуют поверхность глубокой бороздки, с которой, как предполагают, связываются сигнальные последовательности.
Слева представлен вид сбоку; справа — вид сверху.
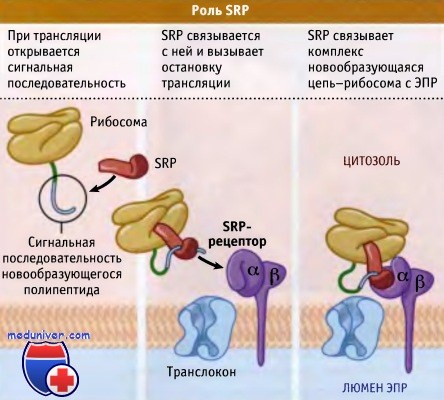
Сразу после выхода сигнальной последовательности с рибосом с ней связывается SRP,
происходит остановка трансляции, и комплекс рибосома-новообразующаяся цепь белка
связывается с мембраной ЭПР посредством взаимодействия с рецептором SRP.
SRP и ее рецептор связывают ГТФ,
высвобождая сигнальную последовательность и способствуя ее проникновению в канал транслокации.
После этого гидролиз ГТФ приводит к диссоциации SRP и рецептора.