• SRP связываются с сигнальными последовательностями
• Связывание SRP с сигнальной последовательностью замедляет трансляцию. Поэтому новообразующийся белок переносится на ЭПР в состоянии незавершенного синтеза и не обладая нативной структурой
• Структурная подвижность М домена SRP54 позволяет SRP узнавать различные сигнальные последовательности
Каким образом сигнальная последовательность направляет белок на ЭПР для последующей транслокации? Открытие SRP показало, что сигнальные последовательности узнаются при специфическом белок-белковом взаимодействии. SRP представляет собой маленькую рибонуклеопротеидную частицу, локализованную в цитозоле, которая содержит шесть полипептидов и небольшую молекулу РНК. Она связывается с сигнальной последовательностью новообразующегося белка, высвобождающегося из рибосомы, и тем самым дает возможность комплексу полипептид-рибосома осуществить взаимодействие с мембраной ЭПР.
Для узнавания сигнальной последовательности требуется только одна субъединица SRP. В соответствии со своей молекулярной массой она называется SRP54, и у многих видов ее структура консервативна, что подчеркивает важность механизма узнавания сигнальных последовательностей.
SRP54 содержит три отдельных домена:
• домен G, связывающий гуанозинтрифосфат (ГТФ) и гидролизующий его до гуанозиндифосфата (ГДФ);
• домен N, представляющий собой N-терминальный домен, который взаимодействует с G-доменом, и
• домен М, который является С-терминальным доменом, содержащим много остатков метионина.
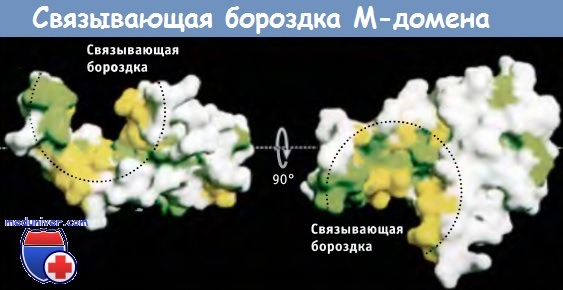
Структура М-домена обеспечивает SRP возможность связываться с различными сигнальными последовательностями, которые ответственны за адресование белков На рисунке ниже представлен М-домен, содержащий несколько а-спиралей, вместе образующих бороздку, в которой и связывается сигнальная последовательность. Изнутри бороздка выстлана остатками метионина. Поскольку боковые цепи метионина обладают гидрофобностью и гибкостью, они действуют подобно гидрофобным ворсинкам и позволяют SRP54 связываться с различными гидрофобными участками, которые находятся в сигнальных последовательностях.
Две другие субъединицы SRP, SRP9 и SRP14, а также 7SPHK присоединяются к рибосоме, снижая скорость элонгации белковой цепи, вероятно, за счет непосредственного вмешательства в процесс связывания трансляционного фактора элонгации. Степень подавления элонгации варьирует от субстрата к субстрату, но во всех случаях подавление снимается только тогда, когда рибосома связывается с ЭПР, и происходит высвобождение SRP. Снижая скорость роста белковой цепи или приостанавливая ее синтез, SRP тем самым обеспечивают доставку новообразующегося полипептида к мембране до того момента, как с рибосомы сойдет большая часть белка.
Это делает маловероятным образование нативной структуры до момента поступления белковой цепи в канал и облегчает процесс переноса. Важность задержки трансляции также подтверждается данными о том, что in vitro белки обычно теряют способность к транлокации, если их узнавание SRP произошло уже после того, как синтезировалась значительная часть полипептидной цепи. Неясно, происходит ли остановка трансляции у всех организмов и даже характерна ли она для всех транслоцируемых белков одного организма. В некоторых случаях доставка новообразующегося белка к мембране происходит достаточно быстро, и тогда остановки трансляции не требуется.
Строение М-домена бактериального гомолога SRP54.
Остатки гидрофобных аминокислот обозначены зеленым и желтым цветом.
Эти остатки образуют поверхность глубокой бороздки, с которой, как предполагают, связываются сигнальные последовательности.
Слева представлен вид сбоку; справа — вид сверху.
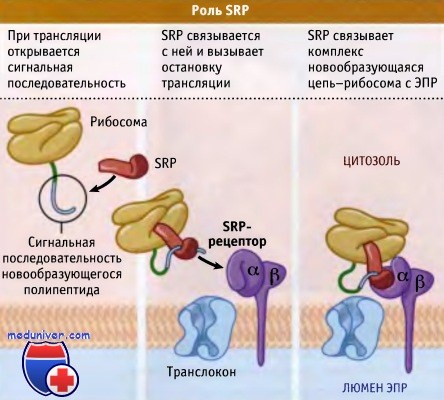
Сразу после выхода сигнальной последовательности с рибосом с ней связывается SRP,
происходит остановка трансляции, и комплекс рибосома-новообразующаяся цепь белка
связывается с мембраной ЭПР посредством взаимодействия с рецептором SRP.