• Транспорт белков через цитоплазматическую мембрану у грамотрицательных и грамположительных бактерий происходит с участием белковых систем Sec и Tat
• Грамотрицательные бактерии также способны транспортировать белки через наружную мембрану
• Для секреции вирулентных факторов патогенные микроорганизмы обладают специальной секреторной системой
Для жизнедеятельности всех клеток необходим безошибочный таргетинг макромолекул. У прокариотов порядка 20% белков транспортируются через цитоплазматическую мембрану. Для всех клеток транспорт белков через биологические мембраны связан с одной и той же проблемой: каким образом можно транспортировать относительно гидрофильную молекулу (белок) через относительно гидрофобную область (мембрану).
Неудивительно, что как про-, так и эукариоты выработали аналогичные системы транспорта белков через мембрану. Обычно аппарат транспорта белков состоит из интегрального мембранного белка или белкового комплекса, образующего канал, через который проходят субстраты. Для протекания этого процесса необходима энергия. Переместившиеся белки связываются с мембраной или выходят во внеклеточное пространство.
Для транспорта большинства белков через цитоплазматическую мембрану у прокариот используются две общие секреторные системы: система Sec, которую также обозначают как общий секреторный путь (GSP), и путь транслокации Tat, который называют «аргининовые близнецы». Наряду с этим, у бактерий обнаружено, по меньшей мере, пять основных классов специализированных секреторных систем, которые обозначаются цифрами от I до V.
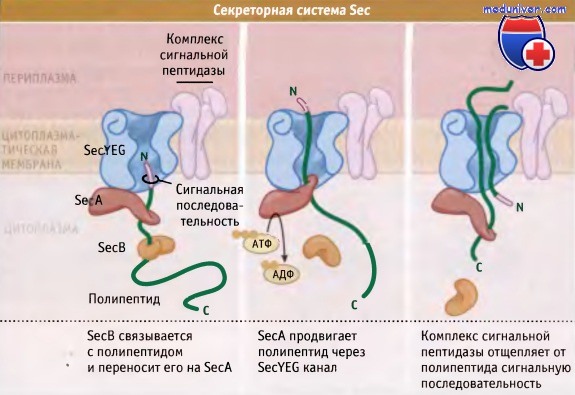
Как показано на рисунке ниже, система Sec состоит из нескольких интегральных мембранных белков, SecY, SecE, SecG и АТФазы, расположенной на периферии мембраны, которая обозначается SecA и обеспечивает энергией процесс транслокации. Для поддержания секретируемого комплекса в несобранном и способном к секреции состоянии для некоторых субстратов Sec у грамотрицательных бактерий необходим цитоплазматический шаперон, SecB.
У грамположительных микроорганизмов гомолог SecB не обнаружен. Все белки, траспортируемые с участием системы Sec, на N-концевом участке содержат сигнальную последовательность, которая направляет их в цитоплазматическую мембрану. Обычно она представлена гидрофобными остатками амиинокислот, однако включает короткий гидрофильный участок, который содержит один или несколько положительно заряженных остатков.
Сигнальные последовательности у грамотрицательных и грамположительных бактерий практически одинаковы, хотя у грамположительных они длиннее, и ближе к N-терминальному участку обогащены положительно заряженными остатками аминокислот. Белки, которые транспортируются с участием системы Sec, могут встраиваться в мембрану, высвобождаться в периплазму, или секретироваться во внеклеточную среду.
Белки общего секреторного пути, или системы Sec,
перемещают новосинтезированные белки через плазматическую мембрану.
Наряду с системой Sec, у бактерий сохранилась еще одна общая секреторная система, Tat. В отличие от Sec, эта система использует протонный градиент для транспорта к клеточной поверхности молекул белков, обладающих нативной структурой, или даже их олигомеров. Система Tat не использует энергию гидролиза АТФ для секреции и не секретирует белковые молекулы, не обладающие нативной структурой. Фактически эта система не использует такие белки в качестве субстратов.
Например, белок PhoA обычно секретируется с участием системы Sec, однако, если в его N-терминальную область включить сигнальную последовательность, специфичную для системы Tat, он становится для нее субстратом. Впрочем, PhoA является субстратом только для Tat, если до секреции в периплазму белок образовал в цитоплазме правильную нативную структуру. Считается поэтому, что прежде чем система Tat узнает субстраты, они должны приобрести правильную нативную структуру.
Белки, входящие в комплекс Tat, включают TatA, В, С и Е. TatABC составляют мембранный комплекс, в то время как TatE является функциональным гомологом TatA. Предполагается, что TatA образует канал, a TatBC комплекс, который связывается с сигнальным пептидом субстрата. Сигальные пептиды, специфичные по отношению к системе Tat, напоминают сигнальные пептиды Sec, однако обладают дополнительными особенностями.
Наиболее характерной их чертой является наличие консервативной последовательности, Ser-Arg-Arg-X-Phe-Leu-Lys, локализованной сразу же после N-терминального участка. Два аргининовых остатка и дали системе название. Почти каждый субстрат Tat содержит два остатка аргинина, хотя некоторые могут содержать только один остаток
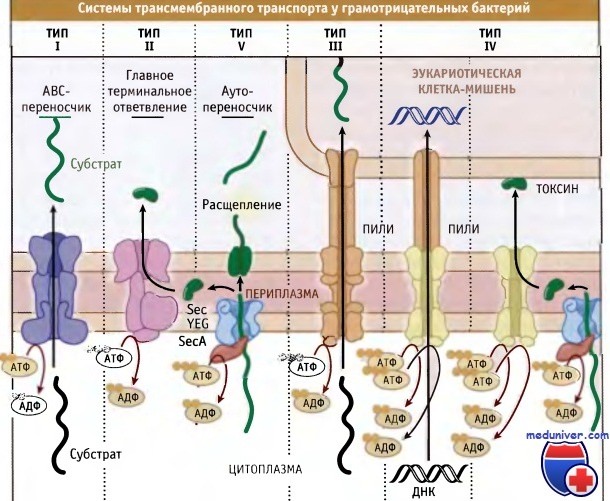
Наряду с общими, у прокариот существуют другие многочисленные системы секреции белков через цитоплазматическую мембрану или, как в случае грамотрицательных микроорганизмов, через цитоплазматическую и наружную мембраны. На рис. 16.26 представлены пять основных групп этих секреторных систем, существующих у грамотрицателных бактерий. Многие из них представляют собой комплексы, состоящие из нескольких субъединиц.
Секреторные системы типа I представляют собой комплекс транспортных белков, связывающий АТФ (АВС) и широко распространенный у бактерий (в клетках эукариот также существует много АВС-переносчиков).Типичная система секреции, принадлежащая к типу I, состоит из двухдоменного белка, локализованного в цитоплазматической мембране. Один из доменов взаимодействует с мембраной, а другой содержит один или два белка, связывающих АТФ.
У грамотрицательных микроорганизмов в состав секреторных систем типа I входит белок наружной мембраны, который транспортирует через нее субстраты. В составе большинства комплексов типа I присутствует дополнительный белок, связанный с цитоплазматической мембраной, который, вероятно, обеспечивает специфичность транспорта. Субстраты системы типа I могут быть самые разные, однако наиболее известны порообразующие токсины и ферменты деградации, секретируемые патогенными бактериями.
У грамотрицательных бактерий секреторная система, относящаяся к типу II (также называемая главное терминальное ответвление GSP), транспортирует через внешнюю мембрану много Sec-зависимых субстратов. Основной характерной чертой главного терминального ответвления является наличие во внешней мембране интегрального белка, который называется секретин. Секретины представляют собой стабильные мультимеры (состоящие из 10-14 субъединиц), которые образуют во внешней мембране канал транслокации.
Обычно для таргетинга и включения во внешнюю мембрану секретинам необходим «белок лоцман». Секретины видны в электронном микросокопе, причем каждый олигомер имеет вид кольцевого комплекса, с центральной порой 10-15 нм в диаметре. Очевидно, такой размер поры достаточно велик для того, чтобы пропустить молекулы белков, которые частично или полностью приобрели нативную структуру.
Система секреции, относящаяся к типу III, исследована очень подробно, поскольку она участвует во взаимодействии патогенных бактерий с организмом хозяина. Субстраты этой системы транспортируются из цитоплазмы бактерий непосредственно в цитоплазму клетки организма хозяина. Таким путем переносятся многие вирулентные факторы, представляющие собой молекулы, которые позволяют патогенным бактериям инфицировать организм и расти в нем. Предполагают, что белки, секретируемые с участием этой системы, в периплазме не образуют промежуточных продуктов.
Вместо этого около 20 белков, участвующих в секреции типа III, образуют комплекс, который пронизывает цитоплазматическую и внешнюю мембраны бактерии и обеспечивает секрецию из клетки «эффекторного» белка. Аналогичная система недавно была описана у грамположительных бактерий.
Системы секреции белков, относящиеся к типу IV, гомологичны системам переноса ДНК. Вначале белковые субстраты этой системы секретируются через цитоплазматическую мембрану посредством системы Sec. Так же как и при функционировании системы III, белки в этом случае секретируются прямо в цитоплазму эукариотической клетки хозяина. Такие системы секреции типа IV, как, например, система переноса Т-ДНК у Agrobacterium tumefaciens, могут переносить в клетки хозяина как белки, так и ДНК. Хотя системы секреции этого типа обнаружены во многих грамотрицательных бактериях, пока для них известны лишь несколько субстатов.
Одним из этих субстратов является коклюшный токсин, который секретируется бактериями Bordetella pertussis с участием системы типа IV. В отличие от переноса Т-ДНК в Agrobacterium, который происходит прямо из цитоплазмы клеток бактерий в цитоплазму клеток хозяина, этот токсин секретируется в два этапа. Токсин проходит через цитоплазматическую мембрану при участии системы Sec, а затем вступает во взаимодействие с системой переноса типа IV и проходит через внешнюю мембрану.
В системах транспорта типа V белок обеспечивает свою собственную секрецию через наружную мембрану. Такие белки называются аутопереносчиками. Вначале они транспортируются в периплазму посредством системы Sec. Например, патогенные бактерии Neisseria используют систему аутопереносчика для секреции протеазы, которая позволяет им избежать атаки иммунной системы организма хозяина. После попадания в периплазму С-концевой участок молекулы белка включается в наружную мембрану и функционирует как переносчик N-концевого участка.
После того как последний достиг клеточной поверхности, он отщепляется от С-концевого участка. Затем протеазный домен отщепляется от поверхности клетки бактерии, а С-концевой остается в составе наружной мембраны. Аутопереносчики, близкие по свойствам к протеазе Neisseria, обнаружены у многих грамотрицательных бактерий.
Основные типы систем трансмембранного транспорта у грамотрицательных бактерий.
Некоторые секреторные системы представляют собой белковые комплексы,
пронизывающие внутреннюю мембрану, периплазму и наружную мембрану.