• In vitro сборка промежуточных филаментов происходит быстро и не требует участия дополнительных факторов
• Во всех белках промежуточных филаментов в центральной части присутствует протяженный а-спирализованный стержневой домен, который необходим для формирования димеров
• Сборка из антипараллельных тетрамеров определяет неполярную природу цитоплазматических промежуточных филаментов
• Сеть промежуточных филаментов является более прочной по сравнению с актиновыми филаментами или микротрубочками, и при нагрузке ее прочность усиливается
В настоящее время на основании данных анализа аминокислотных последовательностей и степени гомологии, а также сведений о биохимических свойствах и кристаллической структуре фрагментов накапливается информация о строении белков промежуточных филаментов.
Первая стадия сборки промежуточных филаментов состоит в образовании суперспирализованных димеров за счет взаимодействия а-спирализованных концевых доменов. Альфа-спираль является наиболее распространенным элементом вторичной структуры белков и впервые была обнаружена именно у белков промежуточных филаментов. У белков промежуточных филаментов а-спирали очень длинные. Для приобретения устойчивости в цитоплазме стержнеобразный домен должен образовать суперспираль с а-спирализованным стержневым доменом другого белка промежуточного филамента.
Необходимость такого взаимодействия объясняется присутствием протяженного гидрофобного участка, который распространяется вдоль а-спирали и служит причиной неустойчивости мономера в водном окружении цитозоля. В а-спирали на каждые семь остатков аминокислот (положения которых в каждой гептаде обозначаются буквами a-g) приходится два витка. В положениях а и d находятся, главным образом, гидрофобные (обычно это лейцин, изолейцин, валин, аланин или метионин) или объемные (фенилаланин или триптофан) остатки.
При образовании суперспирали гидрофобные остатки в положениях а и d в обоих полипептидах оказываются между двумя спиралями. (Эта последовательность а-спирали в белках иногда называется «лейциновая молния».) Структурным элементом 10-нм промежуточного филамента является суперспирализованный димер, в длину достигающий 45 нм. Если такого взаимодействия между белками промежуточных филаментов не происходит, то мономеры скручиваются неправильно и подвергаются протеолизу.
Исследования кератинов помогли прояснить многие детали начальных этапов сборки ранних филаментов, поскольку исследователи могли опираться на закономерности образования димеров тип I/тип II. В клетке мономер кератина неустойчив и начинает деградировать, поэтому для того, чтобы определить, какие особенности структуры способствуют образованию димеров, в клетки дополнительно трансфецируют различные кератиновые конструкции.
В подобных экспериментах было показано, что в клетке мономеры, действительно, оказываются неустойчивыми, и в отличие от гетеродимеров кератинов, гомодимеры не способны образовывать филаменты.
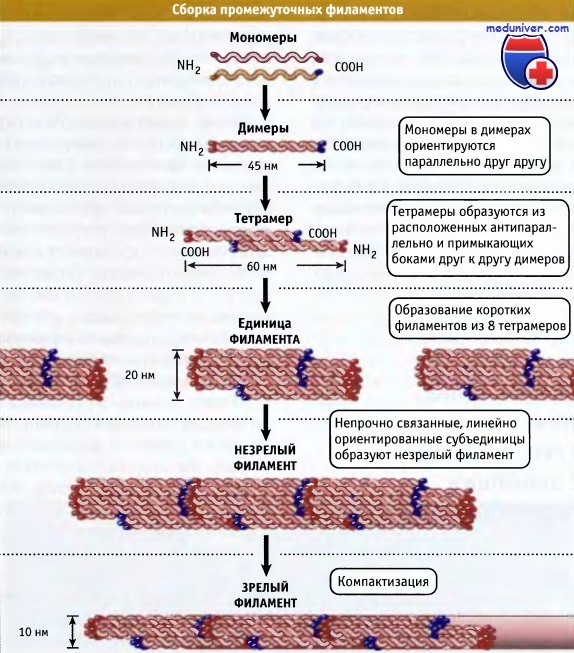
Промежуточные филаменты имеют неполярную природу и представляют собой полифибриллярные полимеры. Это сильно затрудняет изучение процесса их сборки. Почти вся доступная информация получена на основании исследований in vitro и относится только к стержневым доменам. На рисунке ниже представлена общая модель сборки цитоплазматических промежуточных филаментов in vitro. Согласно этой модели, димеры белков промежуточных филаментов цитоплазмы быстро образуют антипараллельные, наполовину смещенные тетрамеры, которые представляют собой минимальные стабильные элементы.
(В отличие от цитоплазматических филаментов, филаменты ядерной ламины собираются в виде параллельных димеров, расположенных ко-нец-в-конец, и соединенные в нити, причем димеры не ассоциируют в тетрамеры латерально.)
Современная модель сборки промежуточных филаментов из мономерных белков.
Образование димеров происходит очень быстро и является необходимым условием предотвращения деградации белка.
По-видимому, тетрамеры являются минимальными по величине агрегатами in vivo. Для простоты неспирализованные домены не показаны.
Последовательности на границе спирализованных и стержневого доменов обозначены красным (со стороны N-концевого участка) и синим цветом (со стороны C-концевого участка).
В клетках сборка в полимерные филаменты происходит, вероятно, быстро. Однако в системах, где процесс может замедляться, он проходит следующие три стадии:
• Быстрая латеральная ассоциация восьми тетрамеров с образованием коротких (60 нм) и толстых (20 нм ширины) «элементарных филаментов» (ULF), после чего
• Ассоциация ULF конец-в-конец в неплотный толстый незрелый филамент, а затем
• Дальнейшая компактизация незрелой структуры с образованием длинного зрелого 10-нм филамента, имеющего гладкую поверхность и среднюю толщину 32 полипептидных цепи.
Эта последовательность стадий сборки in vitro описана для нескольких белков промежуточных филаментов клеток различных организмов, хотя скорость и эффективность протекания отдельных стадий для разных белков варьируют. Предложенная модель сборки согласуется с характером аминокислотных последовательностей в белках промежуточных филаментов: латеральная ассоциация тетрамеров с образованием ULF происходит за счет взаимодействия в стержневом домене заряженных остатков, а ассоциация тетрамеров в филаменты по типу конец-в-конец происходит за счет консервативного мотива, расположенного на границе спирального участка.
Перекрывание пептидов, расположенных на границе спиральных участков, которое происходит по мере сборки субъединиц в филаменты, позволяет объяснить, почему мутации в этих последовательностях влекут за собой тяжелые последствия, и мутации по этим сайтам в белках К5/К14 приводят к сильному поражению кожи при EBS.
Анализ взаимодействий, которые существуют между ближайшими соседями зрелых филаментов, подтверждает наличие гораздо большего числа латеральных взаимодействий, чем в тетрамере. Некоторые из этих взаимодействий могли сформироваться при компактизации, за счет более плотной упаковки субъединиц, продольного сдвига цепей, а также взаимодействий между ними, т. е. когда филамент принимает зрелую конфигурацию.
Дальнейшее формирование филаментов может обеспечиваться взаимодействиями между концевыми доменами, участие которых в упаковке филамента в рамках модели, представленной на рисунке ниже, не учитывается. Исходя из данных структурного анализа, можно предполагать, что головные домены способны взаимодействовать со стержневыми доменами той же или соседней молекулы и что хвостовые домены могут быть более компактными (как в ламинах).
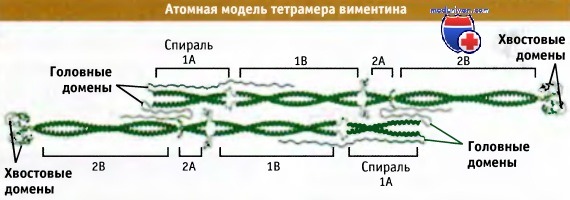
На рисунке ниже представлена модель строения тетрамера белка виментинд, построенная на основании информации, связывающей кристаллическую структуру участка стержневого домена виментина с расчетными структурами. В этой модели можно видеть, насколько протяженный участок занимают закрученные вокруг друг друга а-спирализованные субдомены каждого из двух димеров, изображенные в виде антипараллельных наполовину смещенных структур, т. е. так, как будто они находятся в составе тетрамера.
Атомная модель виментина, белка группы III, представляющего собой гомополимер.
Модель построена на основании данных по кристаллической структуре, имеющихся для отдельных частей молекулы, и гомологичного моделирования ее остальных частей.
Когда белок собирается в филаменты, головные домены, вероятно, ассоциируют со стержневыми в отмеченных областях.
В зависимости от типа промежуточного филамента, димеры представляют собой гомодимеры, или гетеродимеры (как кератины).
О сборке промежуточных филаментов in vivo известно немного. По-видимому, в клетке содержится мало неполимеризованных белков промежуточных филаментов (до 5% простых кератинов, в то время как для актина и тубулина эта цифра составляет 25-50%). Из-за нестабильности длинного а-спирального стержневого домена после синтеза белков должна происходить быстрая сборка филаментов. Возможно также участие шаперонов, которые связываются с белками-мишенями, обычно на промежуточных этапах сборки, и обеспечивают формирование у них правильных пространственных структур.
Некоторые шапероны, такие как, например, небольшие белки теплового шока Hsp27 и а-, b-кристаллины, взаимодействуют с белками промежуточных филаментов после получения клеткой повреждений. Для того чтобы обеспечить формирование нативной структуры, белки промежуточных филаментов должны взаимодействовать друг с другом (т. е. должно происходить быстрое образование суперспирального димера с другим белком с тем, чтобы предотвратить образование неправильной структуры и ее деградацию). Поэтому в роли шаперона для молекулы белка промежуточных филаментов может выступать другой белок промежуточных филаментов.
Наблюдения за характером экспрессии белков в дифференцирующихся клетках позволяют предполагать, что структура филамента может меняться за счет конкурентного замещения предшествующих белков в филаментах вторичными белками. Таким образом, первичные филаменты могут использоваться в качестве матрицы для сборки вторичных филаментов.
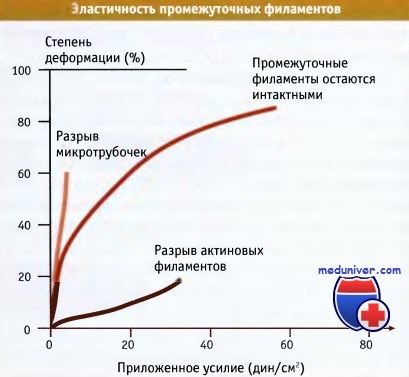
По данным экспериментов in vitro, промежуточные филаменты обладают гораздо большей прочностью на разрыв, чем микротрубочки и актиновые филаменты. Исследования растяжения кератиновых промежуточных филаментов показали их высокую прочность, эластичность, и формирование пластичных необратимых деформаций. Когда in vitro к разным типам промежуточных филаментов применяется механическое воздействие, они проявляют различные биофизические свойства. В отличие от микротрубочек и актиновых филаментов, промежуточные филаменты обнаруживают увеличение устойчивости к деформациям по мере усиления механического воздействия (это свойство называется механическое упрочнение).
Промежуточные филаменты гораздо более устойчивы на разрыв,
чем микротрубочки или актиновые волокна, которые разрываются при небольших нагрузках.
Устойчивость к разрыву измеряли по степени деформации (растяжения),
которую вызывали усилием, приложенным к филаментам, выделенным из клеток.