Механизм полимеризации актина в актиновые филаменты
• В присутствии физиологических концентраций моновалентных и двухвалентных катионов мономерный актин полимеризуется, образуя филаменты
• Актиновые филаменты структурно поляризованы и два их конца неидентичны
В присутствии моновалентных и двухвалентных канонов во внутриклеточной концентрации, мономерный актин полимеризуется с образованием филаментов, диаметр которых составляет 8 нм. Полимеризация носит обратимый характер, и мономеры постоянно добавляются к концу филамента и отщепляются от него. Процесс сборки и разборки обусловливает подвижность клеток, в основе которой лежит процесс полимеризации актина.
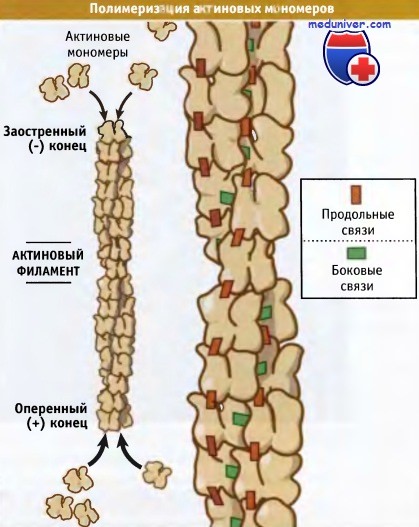
Как показано на рисунке ниже, мономеры полимеризуются, ориентируясь «голова к хвосту», и, таким образом, все субъединицы в филаменте имеют одинаковую ориентацию. Актиновый филамент напоминает двойную нитку бус, закрученную вправо, как показано на рисунке ниже. Это было обнаружено с помощью электронной микроскопии.
Поскольку структура мономеров полярна, филамент также полярен, т. е. два его конца различны. Они называются оперенный и заостренный концы. Полярность филамента играет роль в двухстороннем транспорте и в поляризации клеток.
Конец актинового филамента, который заканчивается субдоменами 1 и 3, представляет собой оперенный конец, а заканчивающийся субдоменами 2 и 4 — заостренный. Эти названия происходят от внешнего вида актиновых филаментов, связанных с миозином. Поскольку актиновые фила-менты спирализованы и поляризованы, в электронном микроскопе миозиновые белки напоминают оперение стрелы. Оперенные и заостренные актиновые филаменты также обозначают как плюс (+) и минус (-) концы соответственно, что согласуется с номенклатурой, принятой для обозначения соответствующих концов микротрубочек.
Каждая актиновая субъединица в составе филамента контактирует с четырьмя соседними субъединицами: одна с каждой стороны той же цепи (продольные контакты) и две на противоположной цепи (латеральные контакты). Эти контакты делают актиновые филаменты очень прочными. Они устойчивы к температурным изменениям и могут расти, достигая длины порядка тысяч субъединиц. Филаменты прочны, но ломаются при резком изгибе. Хотя короткие филаменты (< 5 мкм) не изогнуты, длинные (> 15 мкм) часто искривлены.
Кадры анимации, иллюстрирующие последовательную сборку актиновых мономеров в актиновый филамент.
Актиновые мономеры собираются «голова к хвосту»,
образуя двунитевые поляризованные филаменты.
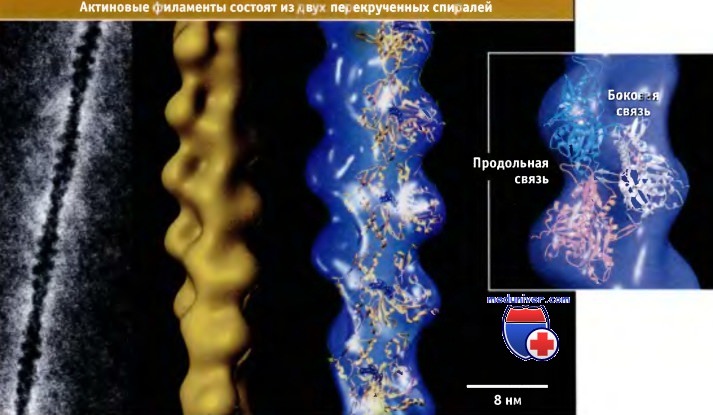
(Слева) Фотография актинового филамента, сделанная с помощью электронного микроскопа.
(В середине) Модель актинового филамента, построенная на основании трехмерной реконструкции электронных микрофотографий.
(Справа) Атомные структуры актиновых мономеров расположены на фоне трехмерной модели и иллюстрируют размещение субъединиц в одной цепи актинового филамента.
• Полимеризация актина de novo представляет собой многоступенчатый процесс, включающий инициацию и элонгацию
• Скорость полимеризации мономера на двух концах филамента различается
• Оперенный конец актинового филамента растет быстрее
Клетки движутся, выдвигая вперед передний край и подтягивая хвостовой отдел. Передние выросты могут представлять собой ламеллоподии или филоподии. Ламеллоподии представляют собой тонкие плоские выросты, окруженные мембраной, содержащие разветвленную сеть актиновых филаментов. Напротив, филоподии выглядят как щупальцы, окруженные мембраной и содержащие пучки актиновых филаментов, расположенных параллельно друг другу. Рост филоподии и ламеллоподии происходит за счет полимеризации актина. Этот процесс мы обсудим здесь подробно.
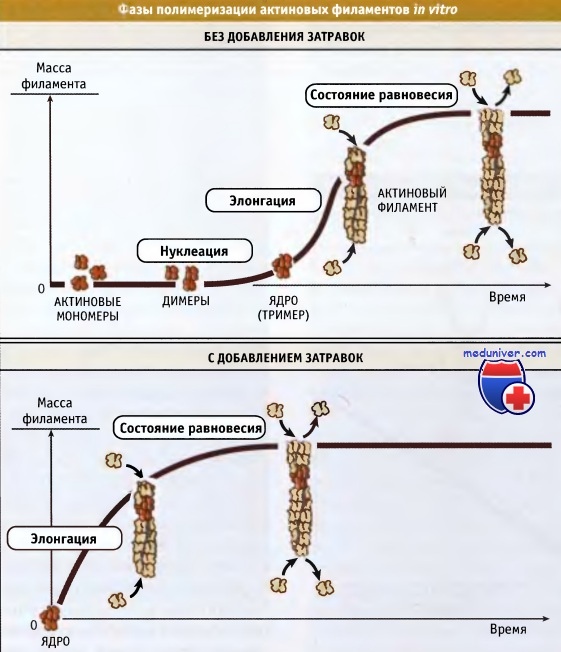
Полимеризация мономеров актина при образовании филаментов представляет собой многоступенчатый процесс. Его можно изучать in vitro, используя очищенный мономерный актин в растворе с низкой ионной силой. Полимеризация инициируется увеличением концентрации соли до физиологического уровня. Процесс спонтанной полимеризации in vitro можно подразделить на три фазы — нуклеация, элонгация, и равновесное состояние.
В фазе нуклеации образуется актиновый олигомер, по свойствам близкий к филаменту. Этот олигомер служит ядром (нуклеусом) для последущей полимеризации актина. Нуклеус представляет собой тример, который является наименьшей единицей, контактирующей с филаментом в продольном и латеральном направлениях. Таким образом, нуклеация происходит в два этапа: образование димера из двух мономеров с последующим присоединением третьей субъединицы. При этом получается тример. Актиновые димеры и тримеры неустойчивы (Kd ~ 100 мкМ-1 мМ) и существуют в очень низких концентрациях. Таким образом, в результате нуклеации при спонтанной полимеризации возникает лаг-период. Лаг-период может отсутствовать, если уже готовы ядра, из которых могут образоваться мономеры (например, если присутствуют преформированные филаменты). В клетке сборка филаментов инициируется белками, связывающимися с актином.
В фазе элонгации происходит быстрый рост филаментов в продольном направлении. Актиновые филаменты растут с концов, а не с боков. Поэтому филаменты различны по длине, но все обладают одинаковой толщиной. По мере полимеризации субъединиц и снижения концентрации свободного мономера, фаза элонгации замедляется.
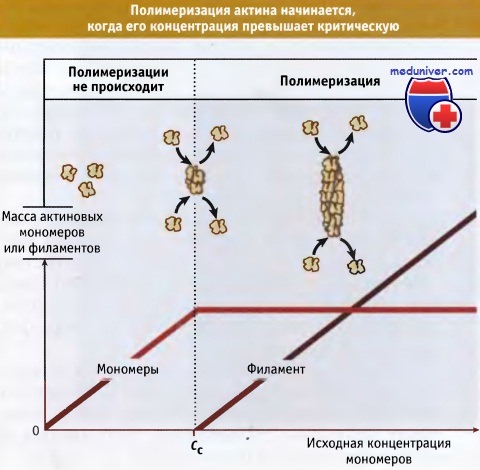
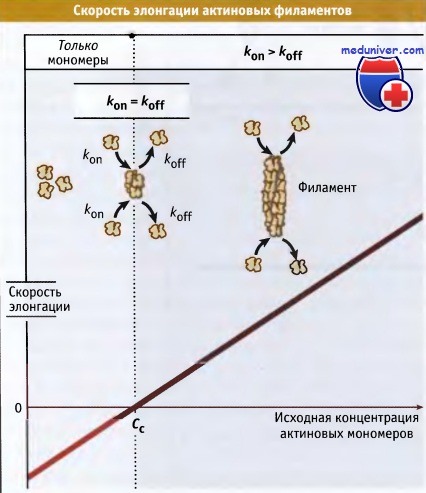
Когда процесс полимеризации находится в стационарном состоянии, то рост филаментов не происходит. Однако наблюдается медленный постоянный обмен между актиновыми субъединицами, находящимися на концах филаментов и в пуле мономеров. Концентрация свободных актиновых мономеров, характерная для стационарного состояния, называется критической концентрацией (Сс).
Этим термином также обозначают концентрацию актиновых субъединиц, которая необходима для образования филаментов (Сс для сборки актина). Когда общая концентрация мономерного актина превышает критическую, происходит образование филаментов. Напротив, если общая концентрация актина ниже критической, он существует только в мономерной форме. При определенном составе раствора критическая концентрация представляет собой постоянную величину, однако она зависит от присутствия регуляторных белков, которые связываются с актиновы-ми мономерами и филаментами.
При концентрации мономеров выше критической происходит элонгация филаментов, скорость которой линейно зависит от концентрации мономеров в растворе. Скорость элонгации индивидуального филамента представляет собой произведение константы скорости добавления мономеров (kon; ед мкМ-1 с-1) и их концентрации. Скорость деполимеризации (koff; ед с-1) не зависит от концентрации мономерного актина. Когда концентрация мономерного актина равняется критической, то скорость элонгации и деполимеризации становится одинаковой, и роста филамента не происходит:
(kon)*(Cc) = koff.
После преобразования получаем:
Cc = koff / kon.
В состоянии равновесия константа диссоциации Kd равняется koff/kon. Поэтому критическая концентрация равняется равновесной константе диссоциации для связывания мономерного актина с концом филамента:
Cc = Kd
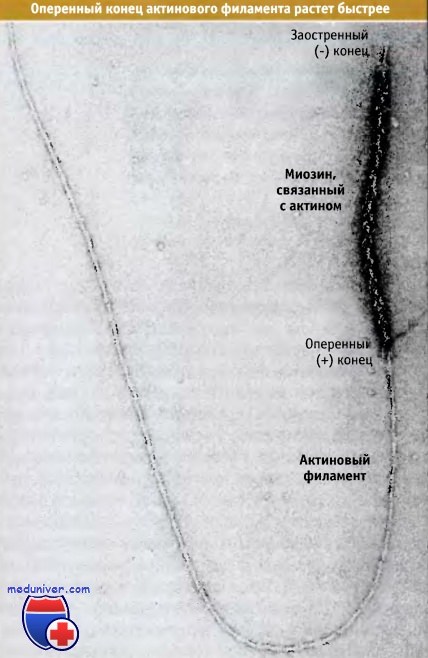
Скорость включения мономера в концы филамента неодинакова. Значения констант скорости элонгации kon и koff (и их отношения, т. е. критической концентрации) зависят от природы нуклеотида, связанного с актином (этот вопрос подробно рассматривается в следующем разделе). В присутствии АТФ мономерный актин связывается с оперенным концом филамента примерно в десять раз быстрее, чем с заостренным концом. Это подтверждается соответствующим экспериментом. В этом эксперименте для нуклеации полимеризации актина использовали короткие актиновые филаменты с присоединенным к ним миозином. На фотографии, сделанной в электронном микроскопе после элонгации филамента, видно, что на оперенном конце полимеризовалось гораздо больше актина, чем на заостренном. Поэтому оперенный конец часто обозначают как быстрорастущий (плюс-конец), а заостренный как медленнорастущий (минус-конец) конец филамента.
Процесс сборки актиновых филаментов, начиная с раствора мономеров, можно подразделить на следующие фазы: нуклеация, элонгация и состояние равновесия.
Фазу нуклеации можно обойти, добавив небольшое количество преформированных филаментов (ядер).
По данным исследований in vitro, сборка акти-новых филаментов начинается,
когда исходная концентрация актиновых мономеров превышает критическую концентрацию, Сс.
По данным экспериментов in vitro, скорость элонгации линейно зависит от концентрации актиновых мономеров.
Результаты эксперимента, демонстрирующего, что полимеризация актина, главным образом, происходит на оперенном конце филамента.
Для нуклеации процесса элонгации использовали филамент, связанный с актином.