• Нуклеосомы могут располагаться в определенных местах. Это происходит вследствие особенностей локальной структуры ДНК либо белков, взаимодействующих с определенными последовательностями
• Наиболее частая причина позиционирования нуклеосом заключается в установлении границы с участием белков, связывающихся с ДНК
• Позиционирование определяет, какие регионы ДНК находятся в линкерной области, и какой стороной она располагается на поверхности нуклеосом
Известно, что in vitro нуклеосомы организуются независимо от первичной структуры ДНК, однако это не означает, что такая же ситуация имеет место in vivo. Всегда ли in vivo определенная последовательность в ДНК находится на определенном месте относительно расположения нуклеосомы? Или же нуклеосомы расположены на ДНК случайным образом, и определенная последовательность может находиться в любом ее участке, например в коровой части в одной копии генома и в линкерной, в другой его копии?
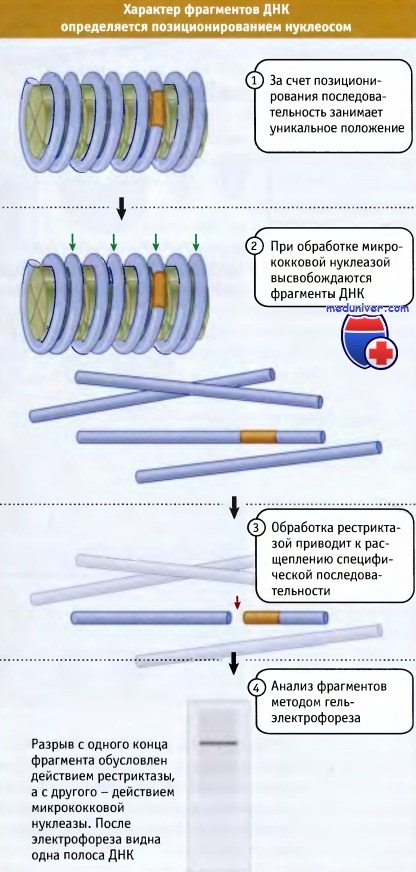
Для ответа на этот вопрос необходимо установить положение определенной последовательности в ДНК, точнее, необходимо установить положение определенного участка ДНК по отношению к нуклеосоме. иллюстрирует принцип используемого метода.
Предположим, что последовательность ДНК организована в нуклеосомы в только одной специфической конфигурации таким образом, что каждый сайт ДНК всегда локализован на нуклеосоме в специфическом положении. Такой тип организации называется позиционированием (или иногда фазированием) нуклеосомы. В серии позиционированных нуклеосом линкерные области ДНК включают уникальные сайты.
Рассмотрим последовательность, находящуюся в пределах одной нуклеосомы. При обработке микрококковой нуклеазой высвобождаются мономерные фрагменты, в том числе фрагмент, содержащий специфическую последовательность. Если этот фрагмент обработать рестриктазой, которая имеет только один сайт узнавания, то он должен разрезаться по специфическому месту. При этом образуются два фрагмента определенной длины.
Фрагменты ДНК, полученные после переваривания микрококковой нуклеазой и рестриктазой, фракционируют методом электрофореза в геле. Для идентификации соответствующего фрагмента в «двойном переваре» используется проба, представляющая собой последовательность на одном конце сайта рестрикции. Такая техника носит название метода гибридизационной концевой метки.
Таким образом, наличие одной полосы свидетельствует о том, что найдено положение сайта рестрикции по отношению к концу нуклеосомной ДНК (по месту ее атаки микрококковой нуклеазой), и нуклеосома позиционирована на определенной последовательности ДНК
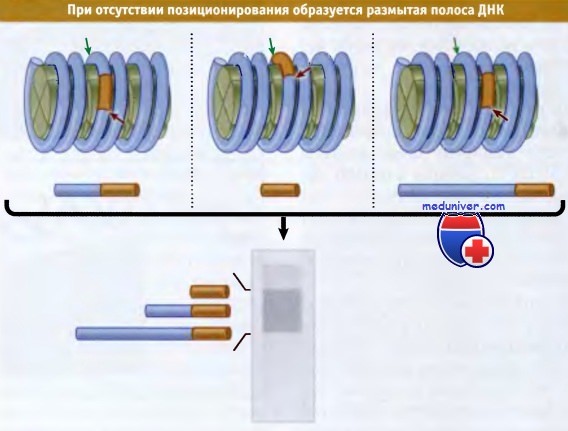
Что происходит, если нуклеосомы не занимают определенного положения? В этом случае в каждой копии генома линкеры содержат различные последовательности ДНК. Таким образом, каждый раз сайт рестрикции находится на разном месте; фактически он может занимать любое местоположение относительно концов мономерной нуклеосомной ДНК Рисунок ниже показывает, что в результате двойного разрыва в ДНК образуется набор фрагментов, варьирующих по длине от наименьшего детектируемого (20 пн) до полного фрагмента мономерной ДНК.
За счет позиционирования нуклеосом сайты рестрикции занимают уникальное положение по отношению к линкерным участкам,
расщепляемым микрококковой нуклеазой.
Позиционирование нуклеосом может осуществляться одним из двух способов:
• Внутренний: каждая нуклеосома специфически формируется в определенной позиции нуклеотидной последовательности. Это меняет нашу точку зрения на нуклеосому, как на субъединицу, способную образовываться при взаимодействии октамера гистонов с любой последовательностью ДНК
• Внешний: первая нуклеосома предпочтительно собирается в определенной позиции региона. Предпочтительная исходная точка для позиционирования нуклеосомы задается регионом, из которого исключаются нуклеосомы. Регион исключения обеспечивает границу, которая ограничивает позиции, доступные для соседних нуклеосом. Затем последовательно может собираться серия нуклеосом, которая характеризуется определенной длиной повтора.
В настоящее время стало очевидным, что расположение октамеров гистонов на ДНК не является случайным по отношению к ее первичной структуре. В одних случаях позиционирование носит внутреннюю природу и определяется структурными характеристиками ДНК В других случаях оно является внешним и зависит от взаимодействия других белков с ДНК и/или с гистонами.
На позиционирование октамеров гистонов влияют некоторые особенности структуры ДНК. Молекуле ДНК присуща тенденция сгибаться преимущественно в одном направлении. Так, АТ-богатые участки локализованы таким образом, что малая бороздка находится со стороны октамера гистонов, а ГЦ-богатые участки расположены так, что она оказывается снаружи.
Длинные участки дА-дТ (> 8 пн) не позиционируются в центральном суперспиральном витке вокруг коровой частицы. Пока еще не представляется возможным учесть все структурные особенности ДНК и предсказать расположение определенной ее последовательности по отношению к нуклеосоме. Также могут играть роль последовательности, которые ответственны за экстремальные процессы, например такие, как исключение нуклеосом, в результате чего могут проявляться граничные эффекты.
Обычно нуклеосомы позиционируются поблизости от границы региона. Если в их строении наблюдается некоторая вариабельность, например длина линкера изменяется, скажем, на 10 пн, то, начиная с первой определенной нуклеосомы, находящейся на границе, специфичность их дальнейшего расположения снижается. В этом случае можно ожидать, что позиционирование будет поддерживаться только для нуклеосом, расположенных относительно близко к границе.
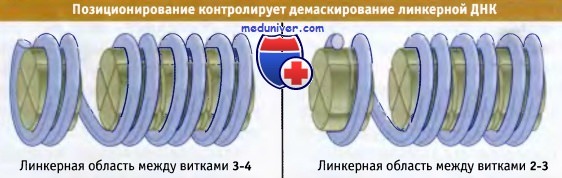
Расположение ДНК в нуклеосоме можно описать с помощью двух моделей. На рисунке ниже показана модель трансляционного позиционирования, которое описывает положение ДНК относительно границы нуклеосомы. В особенности это относится к последовательностям, находящимся в линкерной области. Сдвиг ДНК на 10 пн вызывает образование следующего витка в линкерной области. Таким образом, трансляционное позиционирование определяет, какие регионы ДНК являются более доступными (по крайней мере по отношению к микрококковой нуклеазе).
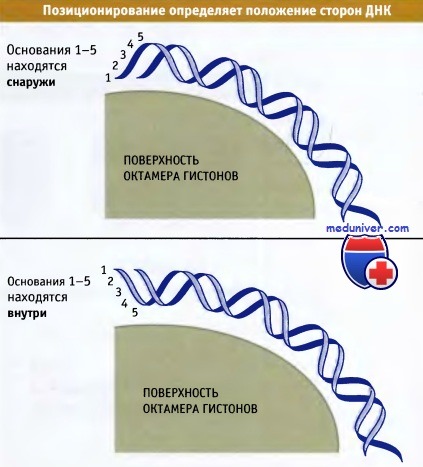
Поскольку ДНК расположена на поверхности октамера гистонов, одна из сторон любой последовательности закрыта белком, а другая остается открытой. В зависимости от расположения на нуклеосоме, сайт ДНК, который должен узнаваться регуляторным белком, может быть доступным или недоступным. Поэтому, важную роль играет точное расположение октамера гистонов по отношению к последовательности ДНК. На рисунке ниже изображена схема ротационного позиционирования двойной спирали по отношению к поверхности коровой частицы. Если ДНК сдвигается на неполный оборот (вообразите, что ДНК вращается по отношению к белковой поверхности), то изменяется степень выхода последовательности за пределы коровой частицы.
Трансляционное и ротационное позиционирование может играть важную роль в обеспечении доступности ДНК. К числу наиболее полно охарактеризованных случаев позиционирования относится специфическое размещение нуклеосом на промоторных участках. Трансляционное позиционирование и/или исключение нуклеосом из определенной последовательности может оказаться необходимым для образования транскрипционного комплекса.
Некоторые регуляторные факторы могут связываться с ДНК лишь в участках, свободных от коровых частиц, поскольку доступность ДНК должна быть максимальной. Это создает границу для трансляционного позиционирования. В других случаях регуляторные факторы могут связываться с ДНК на поверхности нуклеосомы, однако ротационное позиционирование важно для открытия той стороны ДНК, на которой расположены соответствующие контактные точки.
При отсутствии позиционирования нуклеосом сайты рестрикции в различных копиях генома располагаются на разных местах.
При обработке ДНК рестриктазой, атакующей специфические последовательности (выделены красным),
и микрококковой нуклеазой, действующей на межнуклеосомные сайты (выделены зеленым),
образуется смесь разных по размеру фрагментов.
Трансляционное позиционирование отражает линейное положение ДНК относительно октамера гистонов.
Смещение ДНК на 10 пн приводит к смещению последовательностей, находящихся в линкерных областях.
Однако это не влечет за собой изменений ориентации сторон ДНК,
т. е. та сторона ДНК, которая примыкала к гистонам, сохраняет свое положение.
Ротационное позиционирование отражает положение сторон ДНК относительно поверхности нуклеосомы.
Любое перемещение, которое отличается от шага спирали (около 10,2 пн на виток),
приводит к смещению ДНК относительно поверхности гистонового кора.
Последовательности, расположенные внутри, в большей степени защищены от нуклеаз, чем расположенные снаружи.