• Многие нуклеопорины содержат повторы коротких последовательностей, например, GLy-Leu-Phe-GLy, X-Phe-X-Phe-Gly и X-X-Phe-Gly. Предполагают, что в процессе транспорта они взаимодействуют с транспортными факторами
• Некоторые нуклеопорины представляют собой трансмембранные белки, которые привязывают ЯПК к ядерной оболочке
• Идентифицированы все нуклеопорины, входящие в ЯПК дрожжевых клеток
• В процессе митоза происходит разборка и сборка ЯПК
• Некоторые нуклеопорины проявляют свойства динамичности: они способны к быстрой ассоциации с ЯПК и к столь же быстрой диссоциации
Белки, из которых состоят ядерные поровые комплексы (ЯПК), называются нуклеопорины. Наиболее подробно изучены нуклеопорины пекарских дрожжей S. cerevisiae, и мы предполагаем, что в настоящее время уже идентифицированы все дрожжевые нуклеопорины. При идентификации этих белков использовали два методических подхода. В одном из подходов применялся генетический метод, т. е. изучали мутантов, дефектных по процессам ядерного транспорта.
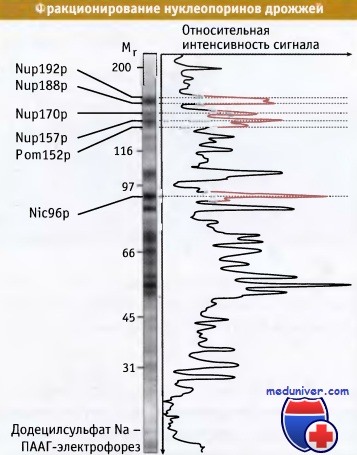
Другой подход представлял собой биохимический и включал выделение индивидуальных ЯПК из препаратов ядерной оболочки. Солюбилизация дрожжевых ЯПК облегчалась отсутствием у дрожжей ядерной ламины. Белки фракционировали с помощью гель-электрофореза, как показано на рисунке ниже, и идентифицировали масс-спектрометрическим методом. При совместном использовании генетического и биохимического подходов удалось идентифицировать примерно тридцать белков.
Были также идентифицированы нуклеопорины Metazoa, в частности клеток амфибий и млекопитающих. Хотя, в этом случае масса ЯПК примерно в два раза превышает массу дрожжевых комплексов ядерных пор, количество находящихся в них нуклеопоринов примерно одинаково. При исследованиях на позвоночных трудно использовать генетические подходы, и поэтому белки ядерных пор были идентифицированы с помощью биохимических и иммунологических методов. Отсутствие ядерной ламины в окончатых мембранах позволило выделить ЯПК из Xenopus.
Многие нуклеопорины характеризуются одной особенностью — наличием множественных повторов, состоящих из коротких последовательностей. Считается, что эти повторы являются сайтами, посредством которых молекулы карго связываются с ЯПК в процессе транспорта. Около 1/3 нуклеопоринов содержат такие последовательности, представленные остатками следующих аминокислот: Gly-Leu-Phe-Gly, X-Phe-X-Phe-Gly, или X-X-Phe-Gly, где X может быть любой аминокислотой.
Поскольку последовательности содержат пару фенилаланин-глицин (Phe-Gly), их часто называют FG-повторы (от англ. Phe-Gly). Последовательности разделяются спейсерами длиной 3-15 аминокислот. Обычно нуклеопорины содержат 10-30 повторов, однако некоторые могут содержать только один или два. Если белок содержит много повторов, они локализуются в одном участке молекулы. Предполагается, что FG-повторы выстилают центральный канал ЯПК, где они выполняют функцию сайтов причаливания для транспортных рецепторов. Еще одна группа нуклеопоринов, содержащих FG-повторы, является компонентами фибрилл цитоплазмы и ядерной корзины.
ЯПК можно выделить из препаратов ядерной оболочки,
а нуклеопорины расфракционировать электрофорезом в геле полиакриламида (ПААГ — полиакриламидный гель).
Показан прокрашенный гель (слева) и результаты его денситометрического анализа (справа).
Помечены некоторые нуклеопорины, присутствующие в больших количествах.
Интересно, что у дрожжевых нуклеопоринов можно удалить много FG-повторов, однако это не сказывается на жизнеспособности клеток. Так, можно удалить все повторы, расположенные в фибриллах и в ядерной корзине, однако в центральном канале возможно удаление лишь нескольких FG-повторов без нарушения жизнедеятельности клетки.
Еще одна особенность многих нуклеопоринов — это присутствие в их структуре мотива, который носит название суперспираль (биспираль). Суперспиральные области представляют собой специфические а-спирали, основная функция которых заключается во взаимодействии с другими а-спиральными доменами с образованием мультимерной структуры. Образование суперспиралей, по-видимому, представляет собой очень важный этап в формировании общей структуры ЯПК.
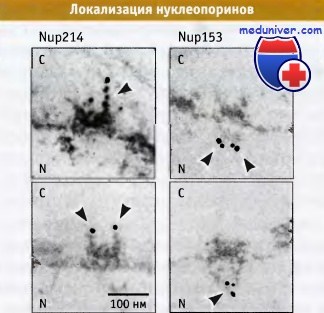
Мы только начинаем понимать, каким образом нуклеопорины организованы в ядерных поровых комплексах (ЯПК). Для выделения субкомплексов ЯПК используются методы биохимического фракционирования, а затем идентифицируются отдельные компоненты и выясняется их общая структурная организация. Этот анализ позволяет выяснить наличие взаимодействия между нуклеопоринами. Локализацию определенного нуклеопорина в ЯПК можно определить с помощью иммуноэлектронной микроскопии. В этом методе для обнаружения специфических нуклеопоринов используются антитела, конъюгированные с частицами золота.
Эти исследования позволяют получить информацию о том, какие нуклеопорины локализуются в цитоплазматических фибриллах, ядерной корзине и в центральной части ЯПК. Большинство нуклеопоринов находится в ядерной и цитоплазматической частях поры, и лишь немного обнаруживается в одной из терминальных структур.
Небольшое количество нуклеопоринов в ядерных поровых комплексах (ЯПК) клеток Metazoa и дрожжей представляют собой трансмембранные белки. К такому выводу позволяют прийти три группы данных:
• Иммуноэлектронная микроскопия показывает, что эти нуклеопорины расположены очень близко к мембране ядерной поры
• Эти нуклеопорины обладают последовательностями аминокислот, достаточно длинными для того, чтобы пронизать липидный бислой
• Эти нуклеопорины находятся в препаратах ядерной оболочки после их обработки солевыми растворами с высокой ионной силой.
После такой обработки остаются только интегральные мембранные белки. Считают, что трансмембранные нуклеопорины способствуют сцеплению ЯПК с ядерной оболочкой.
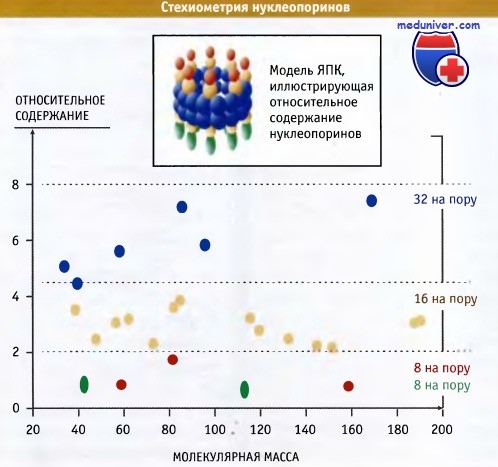
Рисунок выше схематически иллюстрирует относительную представленность нуклеопоринов в различных частях структуры ЯПК дрожжей. Некоторые белки встречаются гораздо чаще других. Поскольку ЯПК характеризуются симметрией 8-го порядка, мы предполагаем, что в одном комплексе находится не менее 8 копий молекулы нуклеопорина. и многие белки присутствуют в 16, 32 или даже большем числе копий. Наиболее редкие нуклеопорины находятся в цитоплазматических фибриллах и ядерной корзине. Значение молекулярной массы и представленность каждого нуклеопорина можно использовать для расчета общей массы дрожжевого ЯПК.
Полученные значения хорошо совпадают с величинами, полученными при измерении размера комплексов с помощью электронной микроскопии, которые составляют 60 млн Да.
Экспериментально показано, что некоторые нуклеопорины прочно связаны друг с другом, в то время как другие проявляют более динамичные свойства. Исследования проводили с использованием метода восстановления флуоресценции после фотоотбеливания (FRAP). Вначале получали клетки млекопитающих, экспрессирующие нуклеопорин, промеченный GFP белком. Затем маленький участок ядерной оболочки подвергался фотоотбеливанию и измеряли кинетику восстановления флуоресценции в отбеленной области. Показано, что примерно половина нуклеопоринов прочно связаны между собой и составляют структурную основу ЯПК.
Остальные белки в течение нескольких часов могут находиться на определенных местах, а небольшое их количество обладают крайне высокой подвижностью, и остаются связанными с ЯПК лишь в продолжении нескольких минут или даже в течение более короткого времени.
В клетках некоторых организмов при митозе происходит разборка ядерной оболочки, и ЯПК диссоциируют на субкомплексы. На поздних стадиях митоза, при образовании ядерной оболочки, ЯПК собираются снова. In vivo процесс диссоциации и повторной сборки занимает менее часа. Используя электронную микроскопию, можно наблюдать разные стадии процесса сборки комплекса. Он состоит из нескольких воспроизводимых шагов. Мы мало знаем о механизме повторной сборки и о том, участвуют ли в нем другие белки, не обнаруженные в комплексе.
Из вновь синтезированных нуклеопоринов в интерфазе также образуются новые ЯПК. Пока мы не знаем, происходит ли начальная сборка de novo и повторная сборка по одному механизму.
В составе ЯПК с помощью метода иммуноэлектронной микроскопии
можно обнаружить индивидуальные нуклеопорины.
Относительное содержание каждого типа нуклеопоринов можно рассчитать на основании данных гель-электрофореза.
Модель иллюстрирует относительное содержание различных нуклеопоринов, локализованных в разных частях ЯПК.
Симметрично расположены только те нуклеопорины, которые находятся в терминальных структурах.