Энергообеспечение мышечного сокращения. Степень перекрытия актиновых и миозиновых нитей
АТФ как источник энергии для сокращения. Химические явления при движении головок миозина. При сокращении мышцы выполняется работа и расходуется энергия. Во время процесса сокращения много молекул АТФ расщепляются с формированием АДФ. Чем больше работа, производимая мышцей, тем больше расщепляется АТФ. Это называют эффектом Фена. При этом, как полагают, происходят следующие явления.
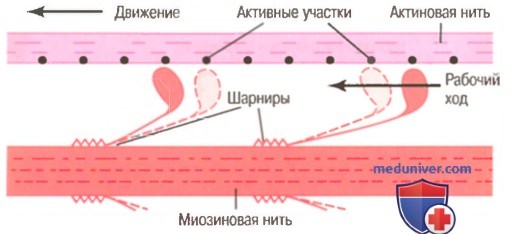
1. До начала сокращения головки поперечных мостиков связываются с АТФ. АТФ-азная активность головки миозина немедленно расщепляет АТФ, но продукты расщепления (АДФ и фосфатный ион) остаются связанными с головкой. В этом состоянии конформация головки такова, что она располагается перпендикулярно нити актина, но еще не прикрепляется к ней.
Храповой механизм сокращения мышцы.
2. Когда тропонин-тропомиозиновый комплекс связывается с ионами кальция, активные участки нити актина открываются, и миозиновые головки соединяются с ними.
3. Связь между головкой поперечного мостика и активным участком на нити актина вызывает конформационное изменение в головке, заставляющее ее наклониться к плечу поперечного мостика. Это обеспечивает рабочий ход для протягивания актиновой нити. Рабочий ход активируется энергией, сохраненной в головке, как во «взведенной» пружине, в виде ее конформационного изменения, происходящего ранее во время расщепления молекулы АТФ.
4. Наклон головки поперечного мостика позволяет освободить АДФ и фосфатный ион, предварительно соединенные с головкой. К месту, от которого выделилась АДФ, прикрепляется новая молекула АТФ, что ведет к отсоединению головки от актина.
5. После отсоединения головки от актина новая молекула АТФ расщепляется, начиная следующий цикл, ведущий к новому рабочему ходу. Это значит, что энергия снова «взводит» головку в перпендикулярное положение, из которого она готова начать новый цикл рабочего хода.
6. Когда «взведенная» головка (с запасом энергии, извлеченной из расщепленной АТФ) связывается с новым активным участком на нити актина, она «разряжается», обеспечивая новый рабочий ход. Таким образом, процесс повторяется снова и снова до тех пор, пока нити актина не притянут Z-диску вплотную к концам миозиновых нитей или пока нагрузка на мышцу не станет слишком большой для осуществления дальнейшей тяги.
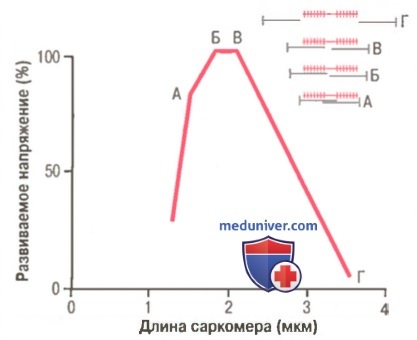
Кривая «длина-напряжение» для одиночного саркомера в условиях изометрического сокращения.
Видно, что максимальная сила сокращения развивается при длине саркомера 2,0-2,2 мкм.
Вверху справа показано относительное расположение актиновых и миозиновых нитей при разной длине саркомера, соответственно точкам от А до Г.
На рисунке демонстрируется влияние длины саркомера и степени перекрытия миозиновых и актиновых нитей на активное напряжение, развиваемое сокращающимся мышечным волокном. Справа черными линиями показаны разные степени перекрытия миозиновых и актиновых нитей при разной длине саркомера. В точке Г на кривой актиновая нить по всей длине оттянута от конца миозиновой нити, и перекрытие между актином и миозином отсутствует. В этом случае напряжение, развиваемое активированной мышцей, равно нулю. Затем по мере укорочения саркомера нить актина начинает перекрывать миозиновую нить, и напряжение постепенно увеличивается до тех пор, пока длина саркомера не уменьшится примерно до 2,2 мкм.
В этом случае актиновая нить перекрывает все поперечные мостики миозиновой нити, но еще не достигает ее центра. При дальнейшем укорочении саркомер сохраняет максимальное на пряжение до тех пор, пока длина саркомера не уменьшится до 2 мкм, что соответствует точке Б на кривой. В этом случае концы двух актиновых нитей начинают перекрывать друг друга в дополнение к перекрытию миозиновых нитей. По мере уменьшения длины саркомера от 2 мкм вплоть до 1,65 мкм, как в точке А, сила сокращения быстро уменьшается. В этой точке два Z-диска саркомера примыкают к концам миозиновых нитей. Если осуществляется сокращение с целью еще большего укорочения саркомера, концы миозиновых нитей изгибаются, и, как видно на рисунке, сила сокращения приближается к нулю, однако вся мышца теперь сжата до своей минимальной длины.
Видео физиология мышц и мышечного сокращения - профессор, д.м.н. П.Е. Умрюхин