Аксонный транспорт. Быстрый аксонный транспорт. Медленный аксонный транспорт.
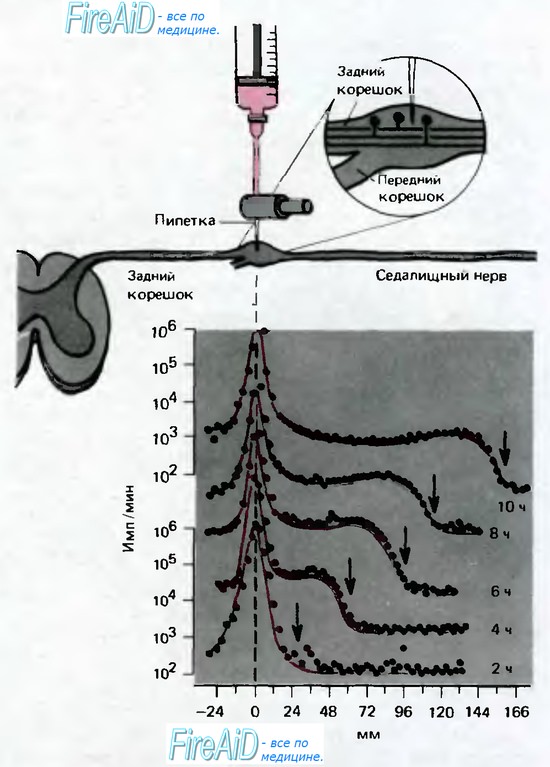
Процессы внутриклеточного транспорта наиболее ярко могут быть продемонстрированы на аксоне нервной клетки. Аксонный транспорт рассматривается здесь подробно, чтобы проиллюстрировать события, которые, вероятно, сходным образом происходят в большинстве клеток. Аксон, диаметр которого составляет всего несколько микронов, может достигать длины одного метра и более, и движение белков путем диффузии от ядра к дисталь-ному концу аксона заняло бы годы. Давно известно, что когда какой-либо из участков аксона подвергается констрикции, часть аксона, расположенная проксимальнее, расширяется. Это выглядит так, как будто в аксоне блокирован центробежный поток. Такой поток быстрый аксонный транспорт-может быть продемонстрирован движением радиоактивных маркеров, как в эксперименте, показанном на рис. 1.14. Лейцин, меченный радиоактивной меткой, инъецировали в ганглий дорсального корешка, и затем со 2-го по 10-й час измеряли радиоактивность в седалищном нерве на расстоянии 166 мм от тел нейронов. За 10 часов пик радиоактивности в месте инъекции менялся незначительно. Но волна радиоактивности распространялась по аксону с постоянной скоростью около 34 мм за 2 ч, или 410 мм/сут. Показано, что во всех нейронах гомойотермных животных быстрый аксонный транспорт осуществляется с такой же скоростью, причем ощутимых различий между тонкими, безмиелиновыми волокнами и наиболее толстыми аксонами, а также между моторными и сенсорными волокнами не наблюдается. Тип радиоактивного маркера также не влияет на скорость быстрого аксонного транспорта; маркерами могут служить разнообразные радиоактивные молекулы, такие, как различные аминокислоты, включающиеся в белки тела нейрона. Если проанализировать периферическую часть нерва, чтобы определить природу переносчиков транспортированной сюда радиоактивности, то такие переносчики обнаруживаются главным образом во фракции белков, но также в составе медиаторов и свободных аминокислот. Зная, что свойства этих веществ различны и особенно различны размеры их молекул, постоянную скорость транспорта мы можем объяснять только общим для всех них транспортным механизмом.
Рис. 1.14. Опыт, демонстрирующий быстрый аксонный транспорт в сенсорных волокнах седалищного нерва кошки. Меченный тритием лейцин вводят в ганглий дорсального корешка и измеряют радиоактивность в ганглии и сенсорных волокнах через 2, 4, 6, 8 и 10 ч после введения (нижняя часть рисунка). По оси абсцисс отложено расстояние от ганглия до участков седалищного нерва, где производят измерение. По оси ординат только для верхней и нижней кривой в логарифмическом масштабе отложена радиоактивность (имп./мин). «Волна» повышенной радиоактивности (стрелки) движется со скоростью 410мм/сут (по [15])
Описанный выше быстрый аксонный транспорт является антероградным, т. е. направленным от тела клетки. Показано, что некоторые вещества движутся от периферии к телу клетки с помощью ретроградного транспорта. Например, ацетилхолинэстераза транспортируется в этом направлении со скоростью в 2 раза меньшей, чем скорость быстрого аксонного транспорта. Маркер, часто используемый в нейроанатомии пероксидаза хрена-также перемещается ретроградным транспортом. Ретроградный транспорт, вероятно, играет важную роль в регуляции белкового синтеза в теле клетки. Через несколько дней после перерезки аксона в теле клетки наблюдается хроматолиз, что свидетельствует о нарушении белкового синтеза. Время, требующееся для хроматолиза, коррелирует с длительностью ретроградного транспорта от места перерезки аксона до тела клетки. Такой результат предполагает и объяснение этого нарушения-нарушается передача с периферии «сигнального вещества», регулирующего белковый синтез.
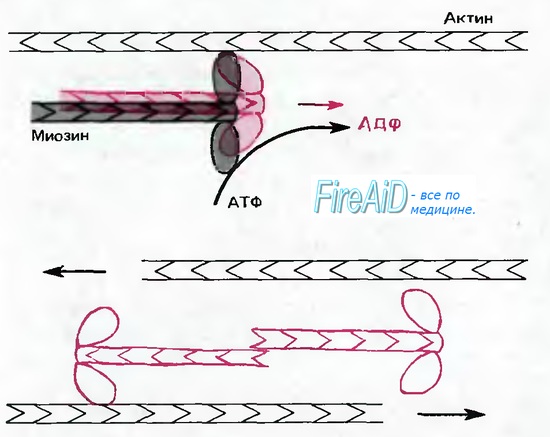
Очевидно, что основными «средствами передвижения», используемыми для быстрого аксонного транспорта, являются везикулы (пузырьки) и орга-неллы, такие, как митохондрии, содержащие вещества, которые нужно транспортировать. Перемещение наиболее крупных везикул или митохондрий можно наблюдать с помощью микроскопа in vivo. Такие частицы совершают короткие быстрые движения в одном из направлений, останавливаются, часто двигаются немного назад или в сторону, снова останавливаются, а затем совершают рывок в основном направлении. 410 мм/сут соответствуют средней скорости антероградного движения приблизительно 5 мкм/с; скорость же каждого отдельного движения должна быть, следовательно, значительно выше, а если учесть размеры органелл, филаментов и микротрубочек, то эти движения действительно очень быстры. Быстрый аксонный транспорт требует значительной концентрации АТФ. Такие яды, как колхицин, разрушающий микротрубочки, также блокируют быстрый аксонный транспорт. Из этого следует, что в рассматриваемом нами транспортном процессе везикулы и орга-неллы движутся вдоль микротрубочек и актиновых филаментов; это движение обеспечивается малыми агрегатами молекул динеина и миозина, действующих, как показано на рис. 1.13, с использованием энергии АТФ.
Быстрый аксонный транспорт может участвовать и в патологических процессах. Некоторые нейротропные вирусы (например, вирусы герпеса или полиомиелита) проникают в аксон на периферии и движутся с помощью ретроградного транспорта к телу нейрона, где размножаются и оказывают свое токсическое действие. Токсин столбняка-белок, который продуцируется бактериями, попадающими в организм при повреждениях кожи, захватывается нервными окончаниями и транспортируется к телу нейрона, где он вызывает характерные мышечные спазмы.
Известны случаи токсического воздействия на сам аксонный транспорт, например воздействие промышленным растворителем акриламидом. Кроме того, полагают, что патогенез авитаминоза «бери-бери» и алкогольной полинейропатии включает нарушение быстрого аксонного транспорта.
Рис. 1.13. Немышечный миозиновый комплекс при определенной ориентации может связываться с актиновыми филаментами различной полярности и, используя энергию АТФ, смещать их относительно друг друга
Помимо быстрого аксонного транспорта в клетке существует и довольно интенсивный медленный аксонный транспорт. Тубулин движется по аксону со скоростью около 1 мм/сут, а актин быстрее-до 5 мм/сут. С этими компонентами цитоскелета мигрируют и другие белки; например, ферменты, по-видимому, связаны с актином или тубулином. Скорости перемещения тубулина и актина примерно согласуются со скоростью роста, обнаруженной для механизма, описанного ранее, когда молекулы включаются в активный конец микротрубочки или мик-рофиламента. Следовательно, этот механизм может лежать в основе медленного аксонного транспорта. Скорость медленного аксонного транспорта примерно соответствует также скорости роста аксона, что, по-видимому, указывает на ограничения, накладываемые структурой цитоскелета на второй процесс.
Завершая данный раздел, следует подчеркнуть, что клетки ни в коем случае не являются статичными структурами, каковыми они кажутся, например, на электронно-микроскопических фотографиях. Плазматическая мембрана и особенно органеллы находятся в постоянном быстром движении и постоянной перестройке; только поэтому они способны функционировать. Далее, это не простые камеры, в которых протекают химические реакции, а высокоорганизованные конгломераты мембран и волокон, в которых реакции протекают в оптимально организованной последовательности.