Передача нервного возбуждения. Участие холинэстеразы в нервном возбуждении
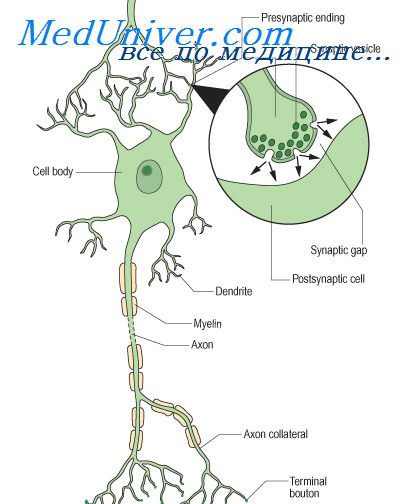
Медиатор нервного возбуждения ацетилхолин синтезируется в нервных клетках из холина и уксусной кислоты при участии фермента холинэстеразы и ацетилкоэнзима А. По последним данным, фермент в нервных клетках содержится не в клеточных органеллах (митохондриях), а локализуется в цитоплазме преимущественно в растворенном виде. ХЭ обладает высоким сродством к мембранам, которое может играть важную роль в синтезе АХ. Медиатор накапливается в окончаниях нервных волокон, где располагается в синаптических пузырьках — везикулах. Каждый пузырек содержит около 10 000 молекул передатчика.
С помощью электронной микроскопии удалось изучить основные структуры синаптических образований в ЦНС и нервно-мышечных синапсах. Установлено, что нервное окончание отделено от синаптической щели пресинаптической мембраной. Ширина синаптической щели — 20—50 нм. За нею расположена постсинаптическая мембрана. В синаптических образованиях гистохимически обнаружены ферменты ХЭ, гидролизующей АХ. Холинэстераза может быть расположена как на пресинаптической, так и на постсинаптической мембране. Это внеклеточная холинэстераза, играющая основную функциональную роль.
В естественных условиях биологические мембраны поляризованы. Величина мембранного потенциала — 50—90 мВ. Когда нервный импульс, распространяясь вдоль аксона, достигает синаптического окончания и деполяризует его, в синапсе происходит быстрое и обратимое увеличение концентрации ионов кальция. Это приводит к перемещению и слиянию части пузырьков с пресинаптической мембраной и в конечном итоге к процессу экзоцитоза — выбросу медиатора в синаптическую щель.
На один нервный импульс в нервном окончании из пузырьков через синаптическую мембрану высвобождается синхронно примерно 150 квантов ацетилхолина. Молекулы медиатора быстро достигают постсинаптической мембраны и взаимодействуют с холинорецептором (ХР), представляющим собой мембранный комплекс белковой природы, прочно встроенный в липопротеидную структуру мембраны.
В состав активного центра холинорецептора входит отрицательно заряженный анионный центр, который реагирует с катионной "головкой" ацетилхолина. Второй участок активного центра — эстерофильный — взаимодействует с карбонильной группой и кислородом эфирной связи молекулы АХ. Конформационные изменения в специфических белках, возникающие под воздействием ничтожных количеств медиатора, могут приводить к "открытию" или "закрытию" каналов к определенным ионам (натрия, калия, кальция, хлора), в результате чего возникает деполяризация этой мембраны, которая регистрируется как потенциал концевой пластинки (ПКП).
Все эти процессы осуществляются в очень короткое время. Для изменения конформации холинорецепторов требуются доли миллисекунды, а для транспорта ионов через мембрану — нескольких миллисекунд. Столь быстрое прекращение этих процессов связано со скоростью ферментативного гидролиза АХ. Возможно, что в прекращении действия АХ некоторая роль принадлежит его диффузии из си-наптической щели и процессам десенситизации — снижению чувствительности рецептора к субстрату. Возникшее в результате нервного импульса изменение проницаемости мембраны и перераспределение ионов восстанавливается с помощью натрий-калиевого насоса, суть которого состоит в активном транспорте ионов против их электрохимических градиентов, что возможно благодаря использованию энергии макроэргических соединений.
Кванты ацетилхолина из отдельных пузырьков могут выделяться спонтанно, без нервного импульса. В ответ на действие одного кванта медиатора появляется слабая (0,2—2 мВ), кратковременная (3—4 мс) деполяризация в виде миниатюрного потенциала концевой пластинки (МПКП). Частота МПКП регулируется пресинаптически и составляет один потенциал в секунду.
При действии антихолинэстеразных веществ может нарушаться каждое из перечисленных выше звеньев цепи синаптической передачи возбуждения. Так, частота выделения квантов существенно возрастает при деполяризации нервных окончаний, что приводит к учащению МПКП. Амплитуда МПКП увеличивается под влиянием антихолинэстеразных веществ, задерживающих разрушение АХ вследствие торможения синаптической ХЭ. Использование микроэлектродной техники позволяет регистрировать увеличение амплитуды и длительности МПКП, а также ПКП при воздействии токсических доз ряда ФОП (ДДВФ, афуган, хостаквик) и некоторых карбаматов — карбофуран, пиримор, диоксикарб.
Установление роли ацетилхолина в процессах передачи нервного возбуждения привлекло внимание исследователей к изучению строения и функции ферментов, названных холинэстеразами, обеспечивающих быстрый гидролиз АХ.